In der vorliegenden Studie wurde versucht, die Rolle von häufig konsumierten künstlichen Süßstoffen auf die Pathogenität von Darmbakterien und die Interaktionen zwischen Darmepithel und Mikrobiota zu untersuchen
Titel im Orginal: Artificial Sweeteners Negatively Regulate Pathogenic Characteristics of Two Model Gut Bacteria, E. coli and E. faecalis
von![]()
![]() Aparna Shil1Biomedical Research Group, School of Life Sciences, East Road, Cambridge CB1 1PT, UK und
Aparna Shil1Biomedical Research Group, School of Life Sciences, East Road, Cambridge CB1 1PT, UK und ![]()
![]() Havovi Chichger2School of Life Sciences, Anglia Ruskin University, Cambridge CB1 1PT, UK – Autor, an den die Korrespondenz gerichtet werden sollte.
Havovi Chichger2School of Life Sciences, Anglia Ruskin University, Cambridge CB1 1PT, UK – Autor, an den die Korrespondenz gerichtet werden sollte.![]()
![]()
Akademischer Redakteur: Miguel Gueimonde
Int. J. Mol. Sci. 2021, 22(10), 5228; https://doi.org/10.3390/ijms22105228
Eingegangen: 19. April 2021 / Überarbeitet: 11. Mai 2021 / Angenommen: 13. Mai 2021 / Veröffentlicht: 15. Mai 2021
Dieser Artikel gehört zur Sektion Molekulare Mikrobiologie
Zusammenfassung
Künstliche Süßstoffe (AS = Artificial Sweeteners) sind synthetische Zuckeraustauschstoffe, die in der Ernährung häufig konsumiert werden. Neuere Studien haben auf erhebliche Gesundheitsrisiken hingewiesen, die den Konsum von AS mit metabolischen Störungen und Störungen der Darmmikrobiota in Verbindung bringen. Trotz dieser Studien gibt es immer noch wenig Daten darüber, wie AS die kommensale Mikrobiota beeinflusst, um Pathogenität zu verursachen.Die Co-Kultur mit humanen Darmepithelzellen zeigt eine Zunahme der Fähigkeit der Modell-Darmbakterien, an das Wirtsepithel zu haften, in es einzudringen und es abzutöten.
1. Einleitung
Es gibt mehr als 100 Billionen Mikroben im Darm, die hundertmal mehr einzigartige Gene kodieren als das menschliche Genom 3Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef]4Wekerle, H. Brain autoimmunity and intestinal microbiota: 100 trillion game changers. Trends Immunol. 2017, 38, 483–497. [Google Scholar] [CrossRef]. Es gibt fortgesetzte, konzertierte Bemühungen, dies weiter zu verstehen, zum Beispiel mit dem METAgenomics of the Human Intestinal Tract Kollaborationsprojekt, das den Beitrag des Mikrobioms bei der Bereitstellung einzigartiger proteinkodierender Gene in einem Umfang untersucht, der 360-mal größer ist als der ihres Wirtes 5Li, J.; Jia, H.; Cai, X.; Zhong, H.; Feng, Q.; Sunagawa, S.; Arumugam, M.; Kultima, J.R.; Prifti, E.; Nielsen, T.; et al. An integrated catalog of reference genes in the human gut microbiome. Nat. Biotechnol. 2014, 32, 834–841. [Google Scholar] [CrossRef]6Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed]. Das Darmmikrobiom befindet sich in unmittelbarer Nähe zum Darmepithel, und als solches sind die Epithelzellen des Darms verschiedenen antigenen Substanzen sowohl aus Nahrungsbestandteilen als auch aus Aktivitäten der Darmmikrobiota ausgesetzt. In der Tat wurde die Bedeutung der kommensalen Mikrobiota für die Funktion des Darmepithels bereits bei keimfreien Mäusen nachgewiesen, die eine signifikante Reduktion der Schleimhautschichtdicke und der antimikrobiellen Produktspiegel aufweisen 7Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, S.; Srinivasan, S.; Ley, R.E.; Gewirtz, A.T. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 2015, 519, 92–96. [Google Scholar] [CrossRef] [PubMed]8Petersson, J.; Schreiber, O.; Hansson, G.C.; Gendler, S.J.; Velcich, A.; Lundberg, J.O.; Roos, S.; Holm, L.; Philipson, M. Importance and regulation of the colonic mucus barrier in a mouse model of colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, 2. [Google Scholar] [CrossRef] [PubMed] 9Vaishnava, S.; Behrendt, C.L.; Ismail, A.S.; Eckmann, L.; Hooper, L.V. Paneth cells directly sense gut commensals and maintain homeostasis at the intestinal host-microbial interface. Proc. Natl. Acad. Sci. USA 2008, 10552, 20858–20863. [Google Scholar] [CrossRef] [PubMed]. Es wurde auch gezeigt, dass mikrobielle Metaboliten aus der Darmmikrobiota, wie kurzkettige Fettsäuren (SCFA), Indol und Laktat, die Funktion der Darmbarriere regulieren 10Kelly, C.J.; Zheng, L.; Campbell, E.L.; Saeedi, B.; Scholz, C.C.; Bayless, A.J.; Wilson, K.E.; Glover, L.E.; Kominsky, D.J.; Magnuson, A.; et al. Crosstalk between microbiota-derived short-chain fatty acids and intestinal epithelial HIF augments tissue barrier function. Cell Host Microbe 2015, 175, 662–671. [Google Scholar] [CrossRef] [PubMed]11Iraporda, C.; Errea, A.; Romanin, D.E.; Cayet, D.; Pereyra, E.; Pignataro, O.; Sirard, J.C.; Garrote, G.L.; Abraham, A.G.; Rumbo, M. Lactate and short chain fatty acids produced by microbial fermentation downregulate proinflammatory responses in intestinal epithelial cells and myeloid cells. Immunobiology 2015, 22010, 1161–1169. [Google Scholar] [CrossRef]. Höhere SCFA-Konzentrationen wurden mit erhöhter Darmdysbiose und Darmpermeabilität in Verbindung gebracht und sind Indikatoren für verschiedene Stoffwechselerkrankungen, während eine verbesserte mikrobielle Diversität eine entgegengesetzte Korrelation mit diesen Ergebnissen zeigte 12de la Cuesta-Zuluaga, J.; Mueller, N.T.; Alvarez-Quintero, R.; Velasquez-Meija, E.P.; Sierra, J.A.; Corrales-Agudelo, V.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Higher fecal short-chain fatty acid levels are associated with gut microbiome dysbiosis, obesity, hypertension and cardiometabolic disease risk factors. Nutrients 2018, 11, 51. [Google Scholar] [CrossRef]. Daher spielen die Darmmikrobiota und die damit verbundenen Stoffwechselprodukte eine bedeutende Rolle für die Darmgesundheit und das Wohlbefinden des Wirts.
Es besteht eine symbiotische Beziehung zwischen Mikrobiota und Wirt, bei der Stoffwechselprodukte der Darmmikrobiota, wie z. B. Vitamine der B-Gruppe und Vitamin K, eine wesentliche Unterstützung für das menschliche Wachstum und die Entwicklung darstellen 13LeBlanc, J.G.; Milani, C.; de Giori, G.S.; Sesma, F.; van Sinderen, D.; Ventura, M. Bacteria as vitamin suppliers to their host: A gut microbiota perspective. Curr. Opin. Biotechnol. 2013, 24, 160–168. [Google Scholar] [CrossRef].
Studien haben gezeigt, dass eine Ernährung, die arm an Fett und tierischem Eiweiß, aber reich an komplexen Kohlenhydraten ist, dem Wirt ein vielfältigeres und damit „gesünderes“ Darmmikrobiom beschert
In den letzten Jahren sind künstliche Süßstoffe als nichtkalorischer Zusatzstoff zum Süßen von Lebensmitteln und Getränken populär geworden. Künstliche Süßstoffe, wie Sucralose und Aspartam, sorgen für den süßen Geschmack in kalorienarmen Lebensmitteln, was ihre Beliebtheit weltweit erhöht hat 22Gardner, C.; Wylie-Rosett, J.; Gidding, S.S.; Sen, L.M.; Johnson, R.K.; Reader, D.; Lichtenstein, A.H.; Physical Activity and Metabolism American Heart Association Nutrition Committee of the Council on Nutrition and American Diabetes Association. Nonnutritive sweeteners: Current use and health perspectives: A scientific statement from the american heart association and the american diabetes association. Diabetes Care 2012, 35, 1798–1808. [Google Scholar] [CrossRef]. Da diese Süßstoffe billig und leicht verfügbar sind und zu einem verbesserten Lebensmittelgeschmack führen, wurden sie in viele Lebensmittel und Getränke sowie in pharmazeutische Produkte aufgenommen. Epidemiologische Studien haben die vorteilhafte Rolle von Süßstoffen bei der Gewichtsabnahme und bei Menschen, die an Glukoseintoleranz und Diabetes mellitus Typ 2 leiden, belegt 23Gardner, C.; Wylie-Rosett, J.; Gidding, S.S.; Sen, L.M.; Johnson, R.K.; Reader, D.; Lichtenstein, A.H.; Physical Activity and Metabolism American Heart Association Nutrition Committee of the Council on Nutrition and American Diabetes Association. Nonnutritive sweeteners: Current use and health perspectives: A scientific statement from the american heart association and the american diabetes association. Diabetes Care 2012, 35, 1798–1808. [Google Scholar] [CrossRef]24Blackburn, G.L.; Kanders, B.S.; Lavin, P.T.; Keller, S.D.; Whatley, J. The effect of aspartame as part of a multidisciplinary weight-control program on short- and long-term control of body weight. Am. J. Clin. Nutr. 1997, 65, 409–418. [Google Scholar] [CrossRef]25Suez, J.; Korem, T.; Zilberman-Schapira, G.; Segal, E.; Elinav, E. Non-caloric artificial sweeteners and the microbiome: Findings and challenges. Gut Microbes 2015, 6, 149–155. [Google Scholar] [CrossRef]; es gibt jedoch auch Studien, die gegenteilige Ergebnisse zeigen.
Anhand von Tier- und Humanstudien wurde der Verzehr von künstlichen Süßstoffen mit Bedingungen in Verbindung gebracht, die zur Entwicklung von Stoffwechselkrankheiten führen.
Interessanterweise zeigen unsere jüngsten Studien an Darmepithelzellen, dass die Exposition gegenüber künstlichen Süßstoffen die Apoptose und die Permeabilität des Darmepithels erhöht, was mit einem entzündlichen Darmleck verbunden ist.
Escherichia coli (E.coli) ist das häufigste fakultativ anaerobe, gramnegative Bakterium, das den menschlichen GI-Trakt besiedelt, und ist ein vielseitiges Pathogen, während Enterococcus faecalis (E. faecalis) ein fakultativ anaerober, grampositiver Kokke ist, der den menschlichen GI-Trakt besiedelt 33Mohamed, J.A.; Huang, D.B. Biofilm formation by enterococci. J. Med. Microbiol. 2007, 56, 1581–1588. [Google Scholar] [CrossRef] [PubMed]34Tenaillon, O.; Skurnik, D.; Picard, B.; Denamur, E. The population genetics of commensal Escherichia coli. Nat. Rev. Microbiol. 2010, 8, 207–217. [Google Scholar] [CrossRef] [PubMed]. Sowohl E. coli als auch E. faecalis sind sowohl Kommensalen als auch Krankheitserreger und besiedeln beide unmittelbar nach der Geburt, so dass sie gute Vertreter ihrer jeweiligen Phyla sind 35Jimenez, E.; Marin, M.L.; Martin, R.; Odriozola, J.M.; Olivares, M.; Xaus, J.; Fernandez, L.; Rodriguez, J.M. Is meconium from healthy newborns actually sterile? Res. Microbiol. 2008, 159, 187–193. [Google Scholar] [CrossRef]. Aus technischer Sicht können beide Spezies leicht isoliert, gezüchtet und im Labor gehalten werden und werden häufig als potenzielle Indikatoren für menschliche Fäkalien verwendet. Wir haben daher E. coli und E. faecalis als Modelle verwendet, um unsere Hypothese zu testen, dass künstliche Süßstoffe bei physiologisch erreichbaren Konzentrationen im Dünndarm 36Shil, A.; Olusanya, O.; Ghufoor, Z.; Forson, B.; Marks, J.; Chichger, H. Artificial sweeteners disrupt tight junctions and barrier function in the intestinal epithelium through activation of the sweet taste receptor, T1R3. Nutrients 2020, 12, 1862. [Google Scholar] [CrossRef]37Mace, O.J.; Affleck, J.; Patel, N.; Kellett, G.L. Sweet taste receptors in rat small intestine stimulate glucose absorption through apical GLUT2. J. Physiol. 2007, 582, 379–392. [Google Scholar] [CrossRef] einen negativen Einfluss auf Bakterien im Darmmikrobiom haben. In der Darmumgebung sind sowohl menschliche Epithelzellen als auch die Mikrobiota verschiedenen Konzentrationen von künstlichen Süßstoffen ausgesetzt, wenn sie mit der Nahrung aufgenommen werden. Wir haben versucht, die Wirkung der häufig konsumierten künstlichen Süßstoffe Saccharin, Sucralose und Aspartam auf zwei Modell-Darmbakterien (E. coli und E. faecalis) zu verstehen, um einen Einblick in die potenziell pathogenen Mechanismen zu erhalten, durch die Süßstoffe die Mikrobiota beeinflussen könnten.
2. Ergebnisse
2.1. Nur der künstliche Süßstoff Saccharin beeinflusst das Wachstum von E. coli-Modell-Darmbakterien bei hohen Konzentrationen
Die Wirkung von künstlichen Süßstoffen auf das Wachstum von E. coli und E. faecalis in planktonischer Kultur wurde alle 12 Stunden gemessen, nachdem sie 4 Tage lang unterschiedlichen Konzentrationen von künstlichen Süßstoffen (Saccharin, Sucralose und Aspartam) ausgesetzt waren. Die Experimente mit E. coli zeigten zu keinem Zeitpunkt und bei keiner Konzentration eine signifikante Veränderung des normalisierten Wachstums als Reaktion auf die Exposition mit Sucralose oder Aspartam (Abbildung 1b oder c). Im Gegensatz dazu reduzierte die Exposition mit 1000 µM Saccharin das E. coli-Wachstum zwischen 48-84 h signifikant, jedoch hatten niedrigere Konzentrationen von Saccharin keinen Einfluss auf das E. coli-Wachstum (Abbildung 1a). Experimente mit E. faecalis zeigten zu keinem Zeitpunkt und in keiner Konzentration eine signifikante Wirkung von Saccharin, Sucralose oder Aspartam auf das normalisierte Zellwachstum der Bakterien (Abbildung 1d-f).
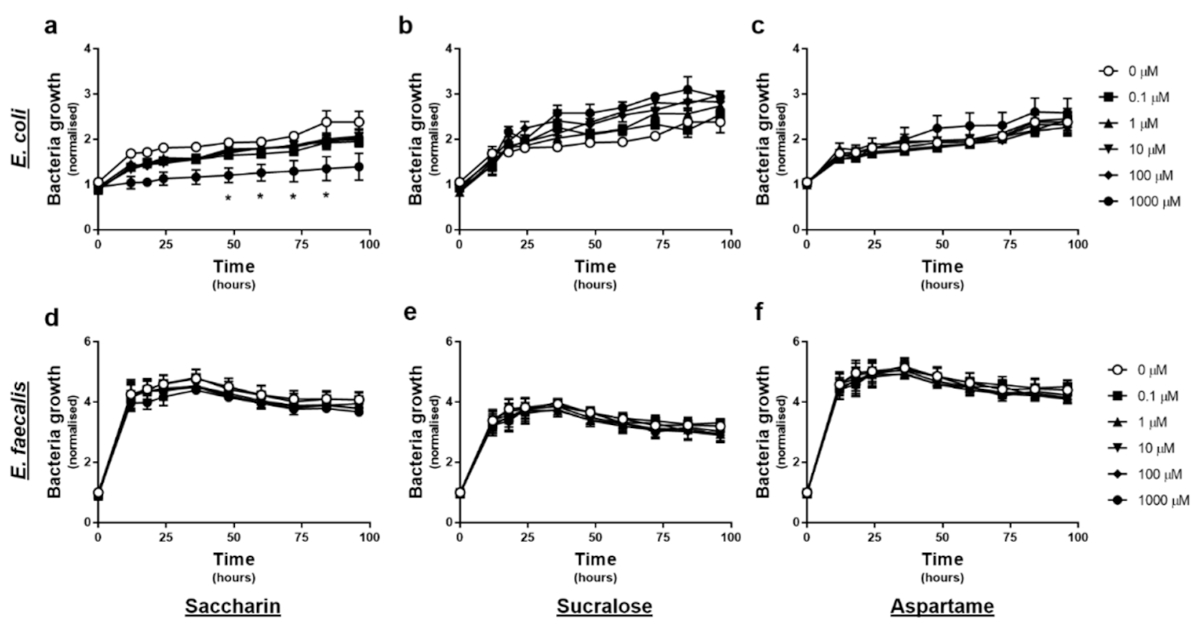
Abbildung 1. Nur der künstliche Süßstoff Saccharin beeinflusst das Wachstum der Darmbakterien des E. coli-Modells bei hohen Konzentrationen. Das Wachstum von E. coli (Tafeln a-c) und E. faecalis (Tafeln d-f) wurde nach Exposition gegenüber den künstlichen Süßstoffen (0 bis 1000 µM) Saccharin (Tafeln a,d), Sucralose (Tafeln b,e) und Aspartam (Tafeln c,f) für bis zu 96 h gemessen. Die Daten sind als Mittelwert ± Standardfehler-Mittelwert (S.E.M.) dargestellt. * p < 0,05 gegenüber 0 µM zum gleichen Zeitpunkt.
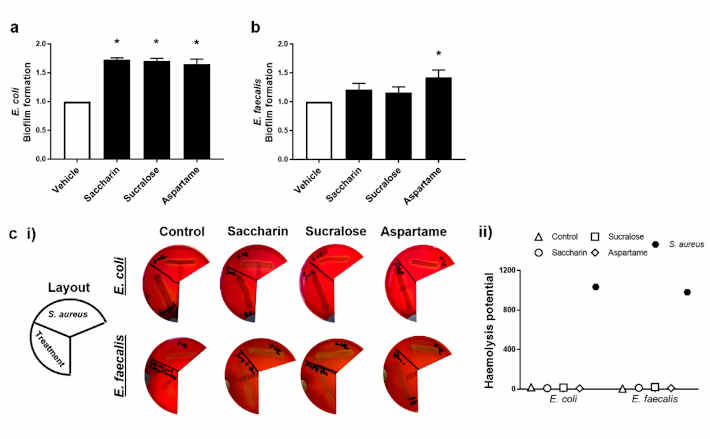
2.2. Künstliche Süßstoffe erhöhen in unterschiedlicher Weise die Biofilmbildung, aber nicht die hämolytische Aktivität, bei den beiden Modell-Darmbakterien
Die Fähigkeit von künstlichen Süßstoffen, die Pathogenität von Modell-Darmbakterien zu regulieren, wurde durch Messung der Biofilmbildung und hämolytischen Aktivität untersucht. Die Experimente wurden mit E. coli und E. faecalis durchgeführt, die Saccharin, Sucralose oder Aspartam in der physiologischen Konzentration von 100 µM ausgesetzt wurden; die Süßstoffe hatten keinen Einfluss auf das Zellwachstum der Bakterien, außer wie bei E. coli und Saccharin bei 1000 µM beobachtet (Abbildung 1a). Die Biofilmbildung von E. coli war im Vergleich zur Vehikelkontrolle signifikant erhöht, wenn die Bakterien den künstlichen Süßstoffen Saccharin, Sucralose und Aspartam ausgesetzt waren (Abbildung 2a). Während die Biofilmbildung von E. faecalis auch mit allen drei künstlichen Süßstoffen erhöht war, verursachte nur die Aspartam-Exposition eine signifikante Erhöhung der Fähigkeit von E. faecalis, einen Biofilm zu bilden (Abbildung 2b). Im Gegensatz dazu zeigte der Hämolyse-Assay, dass die künstlichen Süßstoffe Saccharin, Sucralose und Aspartam keinen Einfluss auf die qualitative (Abbildung 2c (i)) oder semi-quantitative (Abbildung 2c (ii)) Hämolysin-Produktion von E. coli oder E. faecalis hatten, im Gegensatz zur Positivkontrolle, den hämolytischen Bakterien S. aureus.
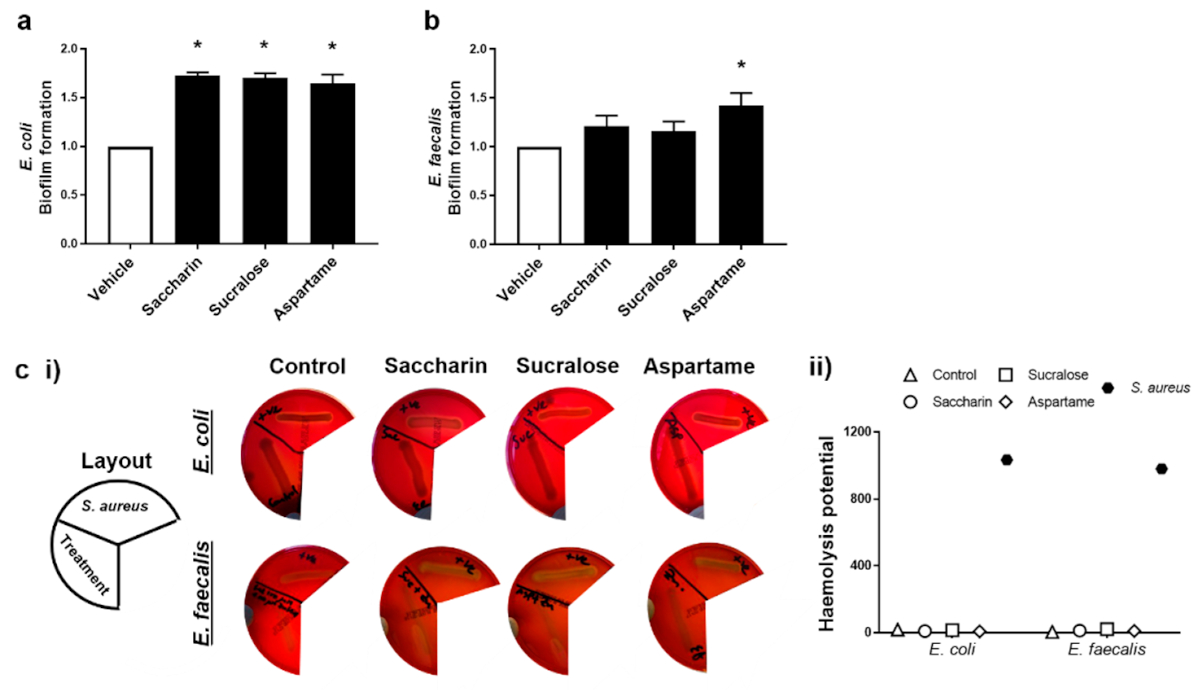
Abbildung 2. Künstliche Süßstoffe erhöhen unterschiedlich die Biofilmbildung, aber nicht die hämolytische Aktivität, in den beiden Modell-Darmbakterien. E. coli (Tafel a) und E. faecalis (Tafel b) wurden den künstlichen Süßstoffen Saccharin, Sucralose und Aspartam (100 µM) für 24 h (Tafel a,b) bzw. 48 h (Tafel c) ausgesetzt und die Biofilmbildung bzw. der Hämolyse-Assay wurde durchgeführt. Für den Hämolyse-Assay wurde S. aureus als Positivkontrolle verwendet, um die Hämolysinfreisetzung (Panel c) anhand von repräsentativen Bildern (Panel c (i)) und Semi-Quantifizierung (Panel c (ii)) der Hämolyse zu zeigen. Spezifische Bakterien für jede Studie sind im unterstrichenen Text enthalten. Die Daten sind als Mittelwert ± S.E.M. für Tafel a und b und als Mittelwert nur für Tafel c dargestellt. * p < 0,05 gegenüber Vehikel für Biofilmbildung.
2.3. Künstliche Süßstoffe stören signifikant die Interaktion zwischen Modell-Darmbakterien und intestinalen Epithelzellen
Um die physiologische Wirkung von künstlichen Süßstoffen auf Modell-Darmbakterien, E. coli und E. faecalis, besser zu verstehen, wurde in den nächsten Studien ein Co-Kultursystem mit der intestinalen Epithelzelllinie, Caco-2-Zellen, verwendet. In den Studien wurde die Wirkung von Saccharin, Sucralose und Aspartam auf die Fähigkeit von Modell-Darmbakterien gemessen, an intestinalen Epithelzellen anzuhaften, in diese einzudringen und sie abzutöten.
Intakte E. coli oder E. faecalis, die mit künstlichen Süßstoffen vorbelastet waren, wurden mit Caco-2-Zellen inkubiert, um die Adhäsionsfähigkeit der Modell-Darmbakterien festzustellen. Alle drei untersuchten künstlichen Süßstoffe, Saccharin, Sucralose und Aspartam, erhöhten signifikant die Adhäsion sowohl von E. coli als auch von E. faecalis an Darmepithelzellen (Abbildung 3a,d). Interessanterweise wurde bei Saccharin (E. coli 2,3 ± 0,4 versus E. faecalis 5,2 ± 2,1), Sucralose (E. coli 2,0 ± 0,3 versus E. faecalis 5,4 ± 1,8) und Aspartam (E. coli 2,9 ± 0,7 versus E. faecalis 6,6 ± 1,9) eine dramatischere Zunahme der bakteriellen Adhäsion an Caco-2-Zellen im Vergleich zu E. coli beobachtet.
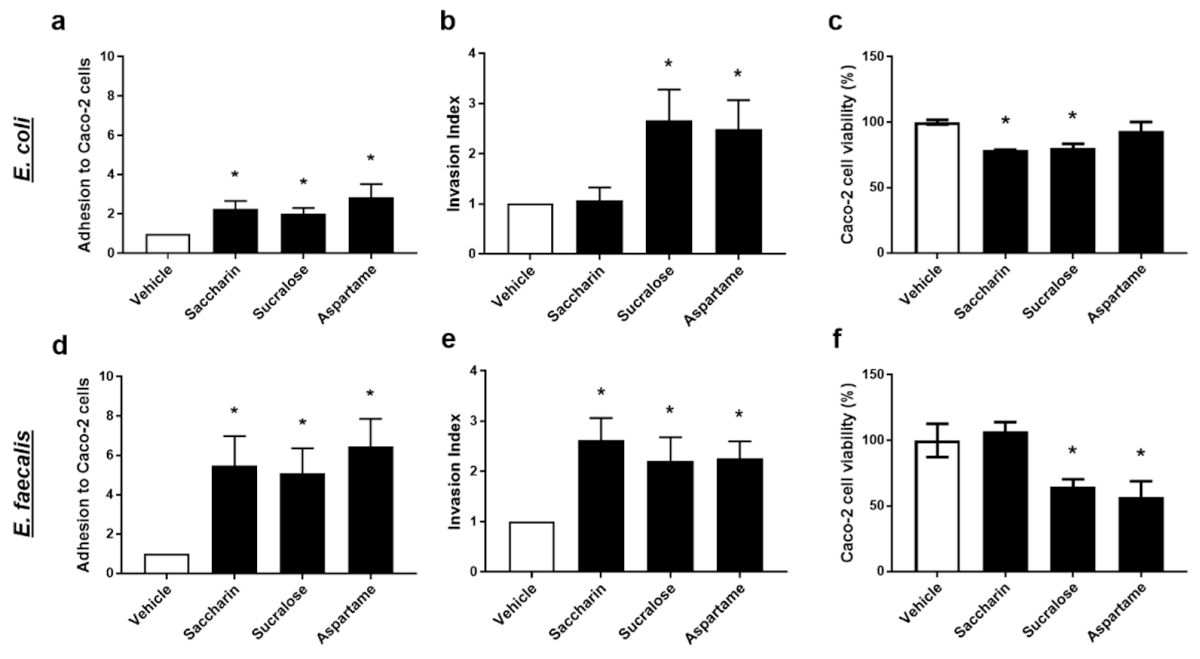
Abbildung 3. Künstliche Süßstoffe stören signifikant die Interaktion zwischen Modell-Darmbakterien und intestinalen Epithelzellen. E. coli (Tafeln a-c) und E. faecalis (Tafeln d-f) wurden 24 h lang künstlichen Süßstoffen, Saccharin, Sucralose und Aspartam (100 µM), ausgesetzt. Die Adhäsion der Bakterien an Caco-2-Zellen (Tafeln a,d) und die Invasion der Bakterien in Caco-2-Zellen (Tafeln b,e) wurden nach 1-stündiger Inkubation mit Caco-2-Zellen gemessen. Die zytotoxischen Effekte der bakteriellen löslichen Faktoren wurden durch 24-stündige Kultivierung des bakteriellen Überstandes mit Caco-2-Zellen (Tafeln c,f) bestimmt. Spezifische Bakterien für jede Studie sind im unterstrichenen Text enthalten. Die Daten sind als Mittelwert ± S.E.M. dargestellt. * p < 0,05 gegenüber Vehikel.
Die Fähigkeit der Modell-Darmbakterien E. coli und E. faecalis, in Caco-2-Zellen einzudringen, wurde nach Exposition mit künstlichen Süßstoffen gemessen. Die Exposition mit Sucralose und Aspartam erhöhte signifikant den Invasionsindex sowohl von E. coli als auch von E. faecalis (Abbildung 3b,e). Im Gegensatz dazu hatte die Inkubation mit Saccharin einen signifikanten Effekt auf die Invasionsfähigkeit von E. faecalis (Abbildung 3e), aber nicht von E. coli (Abbildung 3b). Lösliche bakterielle Faktoren, die von E. coli oder E. faecalis freigesetzt wurden, die künstlichen Süßungsmitteln ausgesetzt waren, wurden mit Caco-2-Zellen inkubiert und die Zytotoxizität wurde gemessen. Lösliche Faktoren, die von E. coli sezerniert wurden, die Saccharin und Sucralose, aber nicht Aspartam ausgesetzt waren, induzierten eine kleine, aber signifikante Reduktion der Lebensfähigkeit von Caco-2 Zellen (Abbildung 3c). Im Gegensatz dazu setzte E. faecalis, das mit Sucralose und Aspartam, aber nicht mit Saccharin exponiert wurde, lösliche Faktoren frei, die die Lebensfähigkeit von Caco-2-Zellen signifikant verringerten (Abbildung 3f).
2.4. Künstliche Süßstoffe beeinflussen Modell-Darmbakterien durch einen geschmackssensitiven Mechanismus
Unsere letzten Experimente versuchten zu verstehen, wie künstliche Süßstoffe die untersuchten pathogenen Funktionen der Modell-Darmbakterien regulieren: Biofilmbildung, Adhäsions- und Invasionsfähigkeit und Zytotoxizität. Aufgrund des Mangels an veröffentlichter Literatur auf diesem Gebiet ist kein spezifischer Geschmacksrezeptor in Bakterien bekannt. Der Pan-Geschmacksinhibitor Zinksulfat wurde verwendet, um das Potenzial eines Geschmackssensormechanismus in E. coli und E. faecalis zu etablieren 38Keast, R.S.; Canty, T.M.; Breslin, P.A. Oral zinc sulfate solutions inhibit sweet taste perception. Chem. Senses 2004, 29, 513–521. [Google Scholar] [CrossRef]. In einer Reihe von Konzentrationen hat Zinksulfat keinen Einfluss auf das Wachstum der beiden Modell-Darmbakterien (Abbildung 4a). Zinksulfat hatte auch keinen Einfluss auf die Biofilmbildung, die Adhäsions- oder Invasionsfähigkeit beider Bakterien in Abwesenheit von Süßungsmitteln (mit Vehikel behandelte E. coli oder E. faecalis) (Abbildung 4b-g). Bei E. coli reduzierte Zinksulfat signifikant die süßstoffinduzierte Biofilmbildung (Abbildung 4b) und die Adhäsionsfähigkeit (Abbildung 4c). Außerdem wurde die Aspartam- und Sucralose-vermittelte Zunahme der Fähigkeit von E. coli, in Caco-2-Zellen einzudringen, durch Zinksulfat abgeschwächt (Abbildung 4d). Bei E. faecalis dämpfte Zinksulfat die durch Aspartam induzierte Biofilmbildung (Abbildung 4e) und alle Süßstoff-induzierten Effekte auf die bakterielle Adhäsion und Invasion von Caco-2-Zellen (Abbildung 4f,g). Die zytotoxische Wirkung auf Caco-2-Zellen, induziert durch Saccharin- und Sucralose-Exposition mit E. coli (Abbildung 3c) und Sucralose- und Aspartam-Exposition mit E. faecalis (Abbildung 3f), wurde ebenfalls durch die Exposition mit Zinksulfat blockiert (Daten nicht gezeigt).
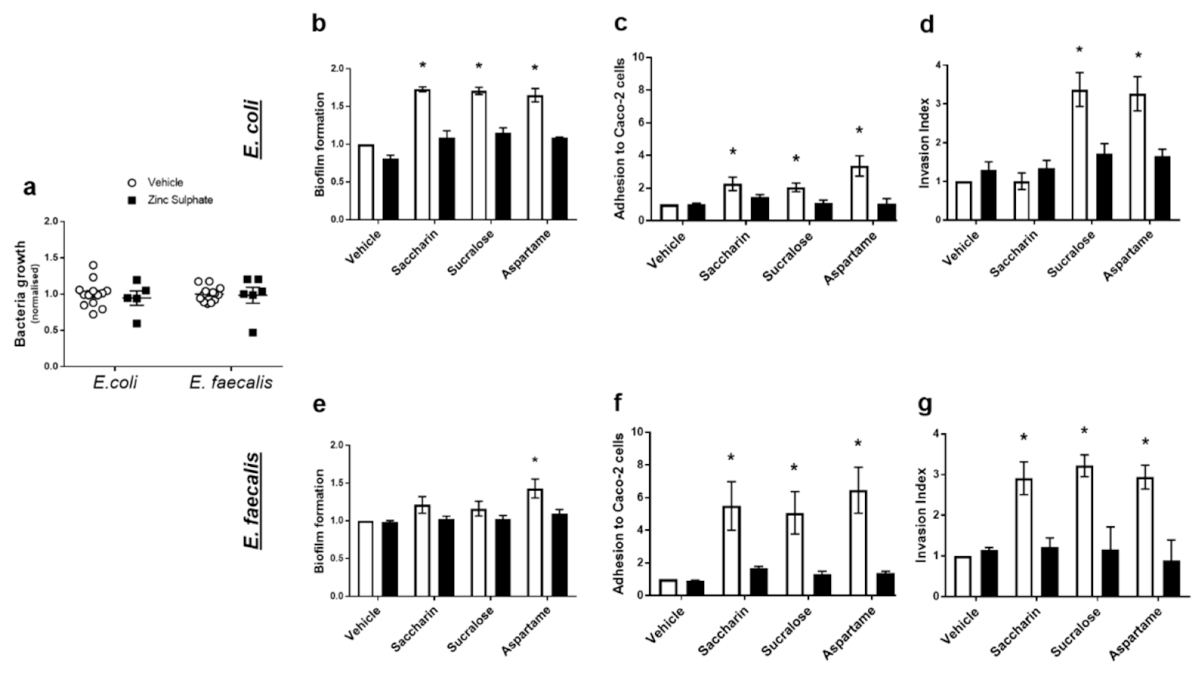
Abbildung 4. Künstliche Süßstoffe beeinflussen Modell-Darmbakterien durch einen Geschmackserkennungsmechanismus. E. coli und E. faecalis wurden 24 h lang Zinksulfat (100 µM) ausgesetzt und das Wachstum wurde gemessen (Tafel a). Alternativ wurden E. coli (Tafeln b-d) und E. faecalis (Tafeln e-g) 24 h lang künstlichen Süßstoffen, Saccharin, Sucralose und Aspartam (100 µM), in Gegenwart oder Abwesenheit von Zinksulfat (100 µM) ausgesetzt. Anschließend wurden Messungen zur Bewertung der Biofilmbildung (Tafeln b,e), der Adhäsion an (Tafeln c,f) und der Invasion von (Tafeln d,g) Caco-2-Zellen durchgeführt. Spezifische Bakterien für jede Studie sind im unterstrichenen Text enthalten. Die Daten sind als Mittelwert ± S.E.M. dargestellt. * p < 0,05 versus Vehikel für Süßstoffe.
Insgesamt zeigen die Ergebnisse, dass die künstlichen Süßstoffe Saccharin, Sucralose und Aspartam die Pathogenität von Modell-Darmbakterien, E. coli und E. faecalis, über einen geschmacksabhängigen Weg erhöhen. Weitere Studien sind erforderlich, um die molekularen Mechanismen zu verstehen, die für diese pathogenen Effekte verantwortlich sind, mit dem Ziel, die negativen Auswirkungen von Süßstoffen auf die Darmgesundheit zu reduzieren.
3. Diskussion
Künstliche Süßstoffe werden in der Ernährung häufig als Hilfsmittel zur Gewichtsabnahme konsumiert, jedoch gibt es immer noch eine Kontroverse über die potenziellen Vorteile oder schädlichen Auswirkungen dieser Verbindungen auf die Darmgesundheit. Frühere Studien haben den Einfluss von Süßstoffen auf die Diversität der Darmmikrobiota gezeigt39Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]40Frankenfeld, C.L.; Sikaroodi, M.; Lamb, E.; Shoemaker, S.; Gillevet, P.M. High-intensity sweetener consumption and gut microbiome content and predicted gene function in a cross-sectional study of adults in the united states. Ann. Epidemiol. 2015, 25, 736–742. [Google Scholar] [CrossRef]41Suez, J.; Korem, T.; Zilberman-Schapira, G.; Segal, E.; Elinav, E. Non-caloric artificial sweeteners and the microbiome: Findings and challenges. Gut Microbes 2015, 6, 149–155. [Google Scholar] [CrossRef]. Diese Studien deuten auf eine Zunahme von Bakterien wie Enterobacteriaceae hin, zu denen auch E. coli gehört, allerdings wurde dies im gesamten Mikrobiom untersucht und nicht auf der Ebene einzelner Bakterien oder auf molekularer Ebene. In der vorliegenden Studie verwenden wir ein In-vitro-Modell des Darms und der Mikrobiota, um den Einfluss von Süßstoffen auf mögliche Veränderungen der Pathogenität zu untersuchen. Unsere Studien zeigen, dass Saccharin, Sucralose und Aspartam in der physiologischen Konzentration von 100 µM, die in der Ernährung leicht erreicht werden kann42Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]43Shil, A.; Olusanya, O.; Ghufoor, Z.; Forson, B.; Marks, J.; Chichger, H. Artificial sweeteners disrupt tight junctions and barrier function in the intestinal epithelium through activation of the sweet taste receptor, T1R3. Nutrients 2020, 12, 1862. [Google Scholar] [CrossRef], die Biofilmbildung sowie die Fähigkeit von Bakterien, an Darmepithelzellen von Säugetieren anzuhaften, in sie einzudringen und sie abzutöten, differenziell erhöhen. Diese Ergebnisse deuten darauf hin, dass Saccharin, Sucralose und Aspartam alle pathogene Veränderungen in zwei Modell-Darmbakterien, E. coli und E. faecalis, fördern, was die Auswirkungen des Konsums von künstlichen Süßstoffen in der Ernährung auf die Darmgesundheit verschlechtern könnte.
Das bakterielle Wachstum ist eine der am besten untersuchten Eigenschaften des Stoffwechsels. Das planktonische Wachstum von E. coli und E. faecalis zeigte wenig Wirkung durch die Exposition gegenüber künstlichen Süßstoffen in einer Reihe von Konzentrationen. Die Ausnahme ist Saccharin in einer Konzentration von 1 mM, das bakteriostatische Effekte in E. coli von 48 bis 84 h verursachte. Diese Ergebnisse unterscheiden sich von denen in der Literatur, wo Wang et al. feststellten, dass Saccharin, Sucralose und Acesulfam K bakteriostatische Effekte auf E. coli HB101 und K-12 Stämme haben 44Wang, Q.P.; Browman, D.; Herzog, H.; Neely, G.G. Non-nutritive sweeteners possess a bacteriostatic effect and alter gut microbiota in mice. PLoS ONE 2018, 13, e0199080. [Google Scholar] [CrossRef]. Wichtig ist, dass diese Studie mit supraphysiologischen Süßstoffkonzentrationen im Bereich von 30-80 mM durchgeführt wurde, was den Unterschied in den Ergebnissen erklären könnte 45Gardner, C.; Wylie-Rosett, J.; Gidding, S.S.; Sen, L.M.; Johnson, R.K.; Reader, D.; Lichtenstein, A.H.; Physical Activity and Metabolism American Heart Association Nutrition Committee of the Council on Nutrition and American Diabetes Association. Nonnutritive sweeteners: Current use and health perspectives: A scientific statement from the american heart association and the american diabetes association. Diabetes Care 2012, 35, 1798–1808. [Google Scholar] [CrossRef]. Unsere Studien zeigen auch, dass alle drei untersuchten Süßstoffe (Saccharin, Sucralose und Aspartam) die Biofilmbildung von E. coli erhöhten, wobei nur Aspartam die Biofilmbildung von E. faecalis beeinflusste. Der Übergang von Bakterien vom Plankton zum Biofilm erfolgt typischerweise in Abhängigkeit von Umweltbedingungen und physiologischen Faktoren wie Zelldichte, Nährstoffverfügbarkeit und zellulärem Stress. Bakterien, die in Biofilmen wachsen, sind weniger empfindlich gegenüber einer antimikrobiellen Resistenzbehandlung und exprimieren im Vergleich zu planktonischen Zellen wahrscheinlich mehr Virulenzfaktoren und Exotoxine 46Ceri, H.; Olson, M.; Morck, D.; Storey, D.; Read, R.; Olson, B. The MBEC assay system: Multiple equivalent biofilms for antibiotic and biocide susceptibility testing. Methods Enzymol. 2001, 337, 377–385. [Google Scholar] [CrossRef]47Kong, K.F.; Vuong, C.; Otto, M. Staphylococcus quorum sensing in biofilm formation and infection. Int. J. Med. Microbiol. 2006, 296, 133–139. [Google Scholar] [CrossRef] [PubMed]48Wagner, V.E.; Li, L.L.; Isabella, V.M.; Iglewski, B.H. Analysis of the hierarchy of quorum-sensing regulation in Pseudomonas aeruginosa. Anal. Bioanal. Chem. 2007, 387, 469–479. [Google Scholar] [CrossRef] [PubMed]. Diese Virulenzfaktoren könnten mit denen verwandt sein, die eine verstärkte Adhäsion an und Invasion von Caco-2-Zellen und erhöhte zytotoxische Effekte verursachen, wie wir in der vorliegenden Studie beobachtet haben. Es ist jedoch erwähnenswert, dass wir bei den verschiedenen Pathogenitätsmessungen, die für jedes Bakterium durchgeführt wurden, nicht das gleiche Muster der Süßstoffwirkung feststellen konnten. Zum Beispiel stellten wir in Studien mit E. coli fest, dass alle 3 untersuchten Süßstoffe eine Zunahme der Biofilmbildung und Adhäsion an Caco-2-Zellen verursachen. Allerdings beeinflussten nur Sucralose und Aspartam die bakterielle Invasion und lösliche Faktoren von nur mit Saccharin und Sucralose behandelten E. coli verringerten die Lebensfähigkeit von Caco-2-Zellen. Daher haben Süßstoffe unterschiedliche Auswirkungen auf die Pathogenität von E. coli und E. faecalis.
Es gibt eine Reihe von Virulenzfaktoren, die Bakterien nutzen können, um für einen Wirt pathogen zu werden, wie z. B. die Verhinderung der Komplementaktivierung und die Flucht aus Phagosomen. Bei einigen pathogenen E. coli wurde gezeigt, dass sie eine Reihe von Virulenzfaktoren aufweisen, wie z. B. bündelbildende Pilus (BFP), Typ 1 Pili und Cytolysin A (ClyA), um die Adhärenz und Invasion von Wirtszellen zu bewirken, sowie die Produktion von Cytotoxinen, die Wirtszellen abtöten49Hyland, R.M.; Sun, J.; Griener, T.P.; Mulvey, G.L.; Klassen, J.S.; Donnerberg, M.S.; Armstrong, G.D. The bundlin pilin protein of enteropathogenic Escherichia coli is an N-acetyllactosamine-specific lectin. Cell Microbiol. 2008, 10, 177–187. [Google Scholar] [CrossRef] [PubMed]50Oscarsson, J.; Mizunoe, Y.; Li, L.; Lai, X.H.; Wieslander, A.; Uhlin, B.E. Molecular analysis of the cytolytic protein ClyA (SheA) from Escherichia coli. Mol. Microbiol. 1999, 32, 1226–1238. [Google Scholar] [CrossRef] [PubMed]51Saldana, Z.; Erdem, A.L.; Schuller, S.; Okeke, I.N.; Lucas, M.; Sivananthan, A.; Philips, A.D.; Kaper, J.B.; Puente, J.L.; Giron, J.A. The Escherichia coli common pilus and the bundle-forming pilus act in concert during the formation of localized adherence by enteropathogenic E. coli. J. Bacteriol. 2009, 191, 3451–3461. [Google Scholar] [CrossRef]. In ähnlicher Weise zeigt pathogener E. faecalis Adhäsion und Invasion intestinaler Epithelzellen über Pili und Aggregationssubstanzen, wie AsaI und Glykolipide, und Zytotoxizität über sezernierte Faktoren, wie Cytolysin52Huycke, M.M.; Joyce, W.A.; Gilmore, M.S. Enterococcus faecalis cytolysin without effect on the intestinal growth of susceptible enterococci in mice. J. Infect. Dis. 1995, 172, 273–276. [Google Scholar] [CrossRef]53Theilacker, C.; Sanchez-Carballo, P.; Toma, I.; Fabretti, F.; Sava, I.; Kropec, A.; Holst, O.; Huebner, J. Glycolipids are involved in biofilm accumulation and prolonged bacteraemia in Enterococcus faecalis. Mol. Microbiol. 2009, 71, 1055–1069. [Google Scholar] [CrossRef]54Wells, C.L.; Moore, E.A.; Hoag, J.A.; Hirt, H.; Dunny, G.M.; Erlandsen, S.L. Inducible expression of Enterococcus faecalis aggregation substance surface protein facilitates bacterial internalization by cultured enterocytes. Infect. Immun. 2000, 68, 7190–7194. [Google Scholar] [CrossRef]55Sartingen, S.; Rozdzinski, E.; Muscholl-Silberhorn, A.; Marre, R. Aggregation substance increases adherence and internalization, but not translocation, of Enterococcus faecalis through different intestinal epithelial cells in vitro. Infect. Immun. 2000, 68, 6044–6047. [Google Scholar] [CrossRef]. Andere Mechanismen der Pathogenität wurden ebenfalls identifiziert. E. coli kann pathogene Effekte, wie Biofilmbildung, durch yafK- und Fis-Genexpression und α- oder β-Hämolyse, möglicherweise über einen ClyA-vermittelten Weg, ausüben56Ludwig, A.; Bauer, S.; Benz, R.; Bergmann, B.; Goebel, W. Analysis of the SlyA-controlled expression, subcellular localization and pore-forming activity of a 34 kDa haemolysin (ClyA) from Escherichia coli K-12. Mol. Microbiol. 1999, 31, 557–567. [Google Scholar] [CrossRef]57Sheikh, J.; Hicks, S.; Dall’Agnol, M.; Phillips, A.D.; Nataro, J.P. Roles for fis and YafK in biofilm formation by enteroaggregative Escherichia coli. Mol. Microbiol. 2001, 41, 983–997. [Google Scholar] [CrossRef]. In ähnlicher Weise wurden Biofilmbildung und Hämolyseaktivität bei pathogenen E. faecalis durch die xdh- oder Esp-Gene bzw. β-Hämolysin nachgewiesen58Heikens, E.; Bonten, M.J.; Willems, R.J. Enterococcal surface protein esp is important for biofilm formation of Enterococcus faecium E1162. J. Bacteriol. 2007, 189, 8233–8240. [Google Scholar] [CrossRef]59Srivastava, M.; Mallard, C.; Barke, T.; Hancock, L.E.; Self, W.T. A selenium-dependent xanthine dehydrogenase triggers biofilm proliferation in Enterococcus faecalis through oxidant production. J. Bacteriol. 2011, 193, 1643–1652. [Google Scholar] [CrossRef] [PubMed]60Shankar, N.; Baghdayan, A.S.; Gilmore, M.S. Modulation of virulence within a pathogenicity island in vancomycin-resistant Enterococcus faecalis. Nature 2002, 417, 746–750. [Google Scholar] [CrossRef]. Während künstliche Süßstoffe nachweislich die Dysbiose in der Darmmikrobiota beeinflussen, gibt es nur wenige mechanistische Studien, die pathogene Reaktionen einzelner Bakterien auf Süßstoffe zeigen. In der vorliegenden Studie zeigen wir, dass die künstlichen Süßstoffe Saccharin, Sucralose und Aspartam in physiologischen Konzentrationen auf alle diese pathogenen Mechanismen mit Ausnahme der β-Hämolyse wirken. Die Modellbakterien E. coli und E. faecalis sind unter normalen Bedingungen α- bzw. γ-hämolytisch, können aber unter pathogenen Bedingungen in β-Hämolyse übergehen 61Shankar, N.; Baghdayan, A.S.; Gilmore, M.S. Modulation of virulence within a pathogenicity island in vancomycin-resistant Enterococcus faecalis. Nature 2002, 417, 746–750. [Google Scholar] [CrossRef]62Giaffer, M.H.; Holdsworth, C.D.; Duerden, B.I. Virulence properties of Escherichia coli strains isolated from patients with inflammatory bowel disease. Gut 1992, 33, 646–650. [Google Scholar] [CrossRef]. Es ist möglich, dass wir aufgrund der In-vitro-Natur der Studie oder der Verwendung von Laborstämmen der einzelnen Bakterien keine Veränderung der Hämolyse festgestellt haben. In der Tat wurde gezeigt, dass klinische Isolate von E. coli oder E. faecalis hämolytische Gene wie hly und ClyA aufweisen63Ike, Y.; Hashimoto, H.; Clewell, D.B. High incidence of hemolysin production by Enterococcus (streptococcus) faecalis strains associated with human parenteral infections. J. Clin. Microbiol. 1987, 25, 1524–1528. [Google Scholar] [CrossRef] [PubMed]64Mirsepasi-Lauridsen, H.C.; Du, Z.; Struve, C.; Charbon, G.; Karczewski, J.; Krogfelt, K.A.; Petersen, A.M.; Wells, J.M. Secretion of alpha-hemolysin by Escherichia coli disrupts tight junctions in ulcerative colitis patients. Clin. Transl. Gastroenterol. 2016, 7, e149. [Google Scholar] [CrossRef] [PubMed], die wahrscheinlich bei den von uns untersuchten Bakterien fehlen. Es ist auch erwähnenswert, dass die Modell-Darmbakterien 24 h lang künstlichen Süßstoffen ausgesetzt waren, so dass nur die lang anhaltende Reaktion auf den Zusatzstoff erfasst wurde. Da künstliche Süßstoffe jedoch ständig in der Nahrung vorhanden sind, und zwar in einer Reihe von Quellen aus Lebensmitteln, Getränken und Kosmetika, ist es wahrscheinlich, dass das Mikrobiom kontinuierlich exponiert wird und die lang anhaltenden Reaktionen am genauesten zu untersuchen sind. Weitere Studien zu den genetischen Veränderungen der einzelnen Bakterien nach Exposition gegenüber Saccharin, Sucralose und Aspartam könnten ein tieferes molekulares Verständnis der Mechanismen liefern, die ihre Pathogenität regulieren.
Bei Säugetieren werden Süßgeschmacksmoleküle über das Süßgeschmacksrezeptor-Hetero- oder Homo-Dimer T1R2/T1R3 bzw. T1R3/T1R3 erkannt 65Nelson, G.; Hoon, M.A.; Chandrashekar, J.; Zhang, Y.; Ryba, N.J.; Zuker, C.S. Mammalian sweet taste receptors. Cell 2001, 106, 381–390. [Google Scholar] [CrossRef]. Zinksulfat ist ein potenter Inhibitor der Süßgeschmackserkennung, von dem angenommen wird, dass er durch die Bindung des Zink-Ions an T1R2/T1R3 bzw. T1R3/T1R3 und die Veränderung der Konformation des Rezeptors wirkt, um die Interaktion von Süßstoffen mit ihm zu verhindern 66Keast, R.S.; Canty, T.M.; Breslin, P.A. Oral zinc sulfate solutions inhibit sweet taste perception. Chem. Senses 2004, 29, 513–521. [Google Scholar] [CrossRef]. Unsere Studien zeigen, dass Zinksulfat die Wirkung von Süßstoffen auf die Pathogenität sowohl für E. coli als auch für E. faecalis effektiv blockieren kann. Dies deutet auf das Vorhandensein eines Süßgeschmackssensors in Bakterien hin, über den Süßstoffe pathogene Wirkungen auslösen können. Sucralose hat jedoch eine wahrgenommene Süßgeschmacksintensität, die 2-mal höher als bei Saccharin und 3-mal höher als bei Aspartam ist 67Wee, M.; Tan, V.; Forde, C. A comparison of psychophysical dose-response behaviour across 16 sweeteners. Nutrients 2018, 10, 1632. [Google Scholar] [CrossRef] [PubMed]. Unsere Ergebnisse zeigen unterschiedliche Effekte der drei Süßstoffe auf die Biofilmbildung, Adhäsion und Invasion sowie Zytotoxizität bei E. coli und E. faecalis. Daher ist es wahrscheinlich, dass diese Bakterien unabhängig von den traditionellen Geschmackserkennungspfaden reagieren. Während Studien auf das Potenzial für Geruchsreaktionen in Bacillus licheniformis hinweisen, gibt es keine Literatur, die auf das Vorhandensein eines Rezeptors oder Sensors für süßen Geschmack in Bakterien hinweist, der auf künstliche Süßstoffe reagieren könnte 68Nijland, R.; Burgess, J.G. Bacterial olfaction. Biotechnol. J. 2010, 5, 974–977. [Google Scholar] [CrossRef]. Stattdessen gibt es Hinweise darauf, dass Süßstoffe DNA-Schäden in Bakterien verursachen, die bakterielle Mutationsrate dosisabhängig erhöhen oder die ROS-Produktion und Entgiftung sowie die Permeabilität der Zellmembran steigern können 69Bandyopadhyay, A.; Ghoshal, S.; Mukherjee, A. Genotoxicity testing of low-calorie sweeteners: Aspartame, acesulfame-K., and saccharin. Drug Chem. Toxicol. 2008, 31, 447–457. [Google Scholar] [CrossRef] [PubMed]70Qu, Y.; Li, R.; Jiang, M.; Wang, X. Sucralose increases antimicrobial resistance and stimulates recovery of Escherichia coli mutants. Curr. Microbiol. 2017, 74, 885–888. [Google Scholar] [CrossRef] [PubMed]71Rencuzogullari, E.; Tuylu, B.A.; Topaktas, M.; Ila, H.B.; Kayraldiz, A.; Arslan, M.; Diler, S.B. Genotoxicity of aspartame. Drug Chem. Toxicol. 2004, 27, 257–268. [Google Scholar] [CrossRef]72Yu, Z.; Wang, Y.; Lu, J.; Bond, P.L.; Guo, J. Nonnutritive sweeteners can promote the dissemination of antibiotic resistance through conjugative gene transfer. ISME J. 2021. [Google Scholar] [CrossRef] [PubMed]. ROS können die Quorum-Sensing-Fähigkeit von Bakterien modulieren, um ihre Umgebung wahrzunehmen und auf sie zu reagieren 73Frey, R.L.; He, L.; Cui, Y.; Decho, A.W.; Kawaguchi, T.; Fergusonm, P.L.; Ferry, J.L. Reaction of N-acylhomoserine lactones with hydroxyl radicals: Rates, products, and effects on signaling activity. Environ. Sci. Technol. 2010, 44, 7465–7469. [Google Scholar] [CrossRef]74Rothfork, J.M.; Timmins, G.S.; Harris, M.N.; Chen, X.; Lusis, A.J.; Otto, M.; Cheung, A.L.; Gresham, D. Inactivation of a bacterial virulence pheromone by phagocyte-derived oxidants: New role for the NADPH oxidase in host defense. Proc. Natl. Acad. Sci. USA 2004, 101, 13867–13872. [Google Scholar] [CrossRef], daher könnten Süßstoffe die Pathogenität der Modell-Darmbakterien in der vorliegenden Studie über einen ROS-abhängigen Weg beeinflussen.
Es wurde gezeigt, dass Bakterien wie E. faecalis durch die Darmwand translozieren, sich in den Blutstrom verbreiten und eine Septikämie zusammen mit einer Ansammlung in den mesenterialen Lymphknoten, der Leber und der Milz verursachen können75Ghoshal, U.C.; Ghoshal, U.; Jain, M.; Kumar, A.; Aggarwal, R.; Misra, A.; Ayyagari, A.; Naik, S.R. Strongyloides stercoralis infestation associated with septicemia due to intestinal transmural migration of bacteria. J. Gastroenterol. Hepatol. 2002, 17, 1331–1333. [Google Scholar] [CrossRef]76Wells, C.L.; Jechorek, R.P.; Erlandsen, S.L. Evidence for the translocation of Enterococcus faecalis across the mouse intestinal tract. J. Infect. Dis. 1990, 162, 82–90. [Google Scholar] [CrossRef]77Wells, C.L.; Erlandsen, S.L. Localization of translocating Escherichia coli, proteus mirabilis, and Enterococcus faecalis within cecal and colonic tissues of monoassociated mice. Infect. Immun. 1991, 59, 4693–4697. [Google Scholar] [CrossRef]. In der vorliegenden Studie konnten wir zeigen, dass Saccharin, Sucralose und Aspartam die Fähigkeit von Modell-Darmbakterien erhöhen, an intestinalen Epithelzellen anzuhaften und in diese einzudringen, mit Ausnahme von Saccharin, das keinen signifikanten Effekt auf die E. coli-Invasion hat. Darüber hinaus haben wir und andere zuvor den negativen Effekt von künstlichen Süßstoffen, Saccharin, Sucralose und Aspartam, auf die Apoptose und Permeabilität von Darmepithelzellen nachgewiesen78Shil, A.; Olusanya, O.; Ghufoor, Z.; Forson, B.; Marks, J.; Chichger, H. Artificial sweeteners disrupt tight junctions and barrier function in the intestinal epithelium through activation of the sweet taste receptor, T1R3. Nutrients 2020, 12, 1862. [Google Scholar] [CrossRef]79Santos, P.S.; Caria, C.R.P.; Gotardo, E.M.F.; Ribeiro, M.L.; Pedrazzoli, J.; Gambero, A. Artificial sweetener saccharin disrupts intestinal epithelial cells’ barrier function in vitro. Food Funct. 2018, 9, 3815–3822. [Google Scholar] [CrossRef], wodurch die Möglichkeit für Bakterien, das Darmepithel zu durchdringen und eine Septikämie zu verursachen, weiter erhöht wird. Bislang wurden jedoch keine Studien durchgeführt, die den Zusammenhang zwischen dem Konsum von künstlichen Süßstoffen und dem Auftreten von Septikämie untersucht haben.
Weltweit wurden Süßstoffe (Acesulfam, Saccharin und Sucralose) in Abwässern, Oberflächenwasser, Grundwasser und Trinkwassersystemen nachgewiesen80Yang, Y.Y.; Liu, W.R.; Liu, Y.S.; Zhao, J.L.; Zhang, Q.Q.; Zhang, M.; Zhang, J.N.; Jiang, Y.X.; Zhang, L.J.; Ying, G.G. Suitability of pharmaceuticals and personal care products (PPCPs) and artificial sweeteners (ASs) as wastewater indicators in the pearl river delta, south china. Sci. Total Environ. 2017, 590–591, 611–619. [Google Scholar] [CrossRef]81Tran, N.H.; Hu, J.; Li, J.; Ong, S.L. Suitability of artificial sweeteners as indicators of raw wastewater contamination in surface water and groundwater. Water Res. 2014, 48, 443–456. [Google Scholar] [CrossRef]82Tran, N.H.; Gan, J.; Nguyen, V.T.; Chen, H.; You, L.; Duarah, A.; Zhang, L.; Gin, K.Y. Sorption and biodegradation of artificial sweeteners in activated sludge processes. Bioresour. Technol. 2015, 197, 329–338. [Google Scholar] [CrossRef] [PubMed]. Noch besorgniserregender ist, dass künstliche Süßstoffe mit einer erhöhten horizontalen Übertragung von antimikrobiellen Resistenzgenen in Umwelt und Klinik in Verbindung gebracht werden83Qu, Y.; Li, R.; Jiang, M.; Wang, X. Sucralose increases antimicrobial resistance and stimulates recovery of Escherichia coli mutants. Curr. Microbiol. 2017, 74, 885–888. [Google Scholar] [CrossRef] [PubMed]84Yu, Z.; Wang, Y.; Lu, J.; Bond, P.L.; Guo, J. Nonnutritive sweeteners can promote the dissemination of antibiotic resistance through conjugative gene transfer. ISME J. 2021. [Google Scholar] [CrossRef] [PubMed]. Darüber hinaus wird vermutet, dass eine verstärkte Biofilmbildung zu Infektionen im Zusammenhang mit medizinischen Geräten führt und eng mit den antibiotikaresistenten Bakterien verbunden ist, die heute eine weit verbreitete Bedrohung der öffentlichen Gesundheit darstellen85Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]86Lleo, M.; Bonato, B.; Tafi, M.C.; Caburlotto, G.; Benedetti, D.; Canepari, P. Adhesion to medical device materials and biofilm formation capability of some species of enterococci in different physiological states. FEMS Microbiol. Lett. 2007, 274, 232–237. [Google Scholar] [CrossRef] [PubMed]87Mack, D.; Rohde, H.; Harris, L.G.; Davies, A.P.; Horstkotte, M.A.; Knobloch, J.K. Biofilm formation in medical device-related infection. Int. J. Artif. Organs. 2006, 29, 343–359. [Google Scholar] [CrossRef] [PubMed]88Braxton, E.E.; Ehrlich, G.D.; Hall-Stoodley, L.; Stoodley, P.; Veeh, R.; Fux, C.; Hu, F.Z.; Quigley, M.; Post, J.C. Role of biofilms in neurosurgical device-related infections. Neurosurg. Rev. 2005, 28, 249–255. [Google Scholar] [CrossRef]. Das Verständnis der Rolle von Süßstoffen bei der Regulierung der Biofilmbildung sowie anderer pathogener Wirkungen auf das Darmepithel und der antimikrobiellen Resistenz von Bakterien könnte die öffentliche Gesundheit in vielerlei Hinsicht dramatisch beeinflussen.
4. Materialien und Methoden
4.1. Materialien
Enterococcus faecalis (E. faecalis, 19433™) und Escherichia coli (E. coli, 10418) wurden von ATCC (Middlesex, UK) bzw. NCTC (Salisbury, UK) bezogen. Staphylococcus aureus, zur Verwendung als Positivkontrolle für den Hämolyse-Assay, war ein freundliches Geschenk von Dr. Christopher O’Kane (Anglia Ruskin University). Bakterielle Medien und Blutagarplatten wurden von Oxoid (ThermoFisher, Hampshire, UK) bezogen. Für das Bakterienwachstumskurven-Experiment und den Biofilm-Assay wurden sterile, unbehandelte 96-Well-Polystyrolplatten mit flachem Boden von CytoOne (StarLabs, Milton Keynes, UK) erworben. Phosphatgepufferte Kochsalzlösung (PBS) wurde von Gibco (ThermoFisher, Hampshire, UK) bezogen. Humane Kolon-Adenokarzinom-Zellen (Caco-2, 86010202), Eagle’s Minimal Essential Media (EMEM), Zinksulfat, Antibiotika und künstliche Süßstoffe (Saccharin, Sucralose und Aspartam) wurden von Sigma-Aldrich (Dorset, UK) bezogen.
4.2. Bakterien- und Säugetierzellkultur
Bakterielle Zellen wurden aseptisch bei 37 °C auf festen Medien für Einzelkolonien oder in flüssigen Medien unter Schütteln (150 U/min) für Wachstumsmessungen gezüchtet. Brain Heart Infusion Agar und Nutrient Agar wurden als feste, Brain Heart Infusion und Nutrient Broth als flüssige Medien für E. faecalis bzw. E. coli verwendet, wie vom Lieferanten vorgegeben.
Eine humane intestinale Epithelzelllinie, Caco-2-Zellen, wurde für die Interaktionsassays, wie Adhäsions-, Invasions- und Zytotoxizitätsassays, verwendet. Monolayer von Caco-2-Zellen wurden aseptisch in EMEM, ergänzt mit 10 % fötalem Rinderserum und 1 % Antibiotika-Lösung (1 U/mL Penicillin, 1 μg/mL Streptomycin) bei 37 °C und 5 % CO2 gezüchtet.
4.3. Bestimmung der Wachstumskurve
Eine einzelne Bakterienkolonie von E. coli oder E. faecalis wurde aseptisch in Nährstoffbrühe bzw. Hirn-Herz-Infusion beimpft, die mit den künstlichen Süßstoffen Saccharin, Sucralose und Aspartam in Konzentrationen von 0,1 bis 1000 µM oder Vehikel (doppelt destilliertes Wasser (ddH2O)) versetzt war, und bis zu 4 Tage lang wachsen gelassen. Das Wachstum wurde als Absorption bei 600 nm (A600) mit dem VictorTM X3 (Perkin Elmer) aufgezeichnet und die Werte wurden auf 0 µM bei 0 h (als 1) normalisiert.
4.4. Assay zur Biofilmbildung
Die Biofilmbildung von E. coli und E. faecalis wurde nach Exposition gegenüber den künstlichen Süßstoffen Saccharin, Sucralose und Aspartam (100 µM) unter Verwendung des Crystal Violet Biofilm Formation Assay gemessen, wie zuvor beschrieben [68] mit einigen Modifikationen. Eine einzelne Bakterienkolonie wurde in 10 mL des entsprechenden flüssigen Mediums beimpft, das mit Süßstoff oder Vehikel (H2O) in An- oder Abwesenheit von Zinksulfat ergänzt wurde. Die Absorption bei 600 nm wurde auf einem VictorTM X3-Multiplattenlesegerät gemessen, um sicherzustellen, dass die Anzahl der Bakterienzellen gleich ist, und die Kultur wurde über Nacht in flüssige Medien (1:200) übertragen, die mit künstlichen Süßstoffen ergänzt wurden. Nach dem Vortexen wurden 200 µL in sterile 96-Well-Kunststoffplatten überführt und für 48 h bei 37 °C aerob gezüchtet. Der Überstand wurde entfernt, und die Vertiefungen wurden zweimal mit ddH2O gewaschen, um lose assoziierte Bakterien zu entfernen. Jede Vertiefung wurde mit 150 µL 0,1% Gram-Kristallviolett für 20 min bei Raumtemperatur gefärbt. Nach der Färbung wurden die Wells dreimal mit ddH2O gewaschen. Das von den Biofilmen zurückgehaltene Kristallviolett wurde durch Zugabe von 200 µL 30%iger Essigsäure in Lösung gebracht und für 5 min bei 37 °C inkubiert. Die quantitative Analyse der Biofilmbildung erfolgte durch Messung der Absorption bei 600 nm mit dem Multiplate-Reader (VictorTM X3). Die Biofilmbildungseinheit wurde berechnet, indem die Absorption des zurückgehaltenen Kristallviolets durch die Absorption des gesamten Bakterienwachstums geteilt und auf die Kontrolle (als 1) normiert wurde.
4.5. Hämolyse-Assay mit Blutagar-Platten
Eine Blutagarplatte mit 7% Pferdeblut wurde verwendet, um die hämolytischen Eigenschaften von E. coli und E. faecalis nach Exposition gegenüber künstlichen Süßstoffen zu untersuchen. Die Bakterien wurden den künstlichen Süßungsmitteln Saccharin, Sucralose und Aspartam (100 µM) für 24 h in An- und Abwesenheit von Zinksulfat (100 µM) unter Schüttelbedingungen bei 37 °C ausgesetzt. Die Absorption der Bakterien wurde bei 600 nm bestimmt und die Bakterienzahl wurde vor dem Ausplattieren auf Blutagarplatten normalisiert. Die Platten wurden bei 37 °C inkubiert und die Hämolyse wurde nach 24 h zusammen mit Staphylococcus aureus als Positivkontrolle bestimmt 89Wiseman, G.M. The hemolysins of Staphylococcus aureus. Bacteriol. Rev. 1975, 39, 317–344. [Google Scholar] [CrossRef] [PubMed]. Die Bilder der Platten wurden mit dem UVI-Tec Imager (Uvitec Ltd., Cambridge, UK) aufgenommen. Um eine semi-quantitative Analyse des Hämolyse-Assays durchzuführen, wurden die Bilder mit ImageJ 1.51 s (http://imagej.nih.gov/ij, 19. März 2021) ausgewertet. Für jeden Streifen wurde die Fläche der Bakterien und der Hämolysefläche auf den Blutagarplatten in relativen Einheiten gemessen und das hämolytische Potenzial wurde als Hämolysefläche minus Bakterienfläche berechnet.
4.6. Adhäsions-Assay
Die Adhäsion der Modell-Darmbakterien an Caco-2-Zellen nach Exposition mit künstlichem Süßstoff wurde wie zuvor beschrieben 90Darfeuille-Michaud, A.; Aubel, D.; Chauviere, G.; Rich, C.; Bourges, M.; Servin, A.; Joly, B. Adhesion of enterotoxigenic Escherichia coli to the human colon carcinoma cell line caco-2 in culture. Infect. Immun. 1990, 58, 893–902. [Google Scholar] [CrossRef] mit einigen Modifikationen gemessen. Caco-2-Zellen wurden auf Gewebekulturplatten mit 24 Vertiefungen (7,5 × 104 Zellen/Vertiefung) ausgesät und unter befeuchteten Bedingungen (90 %) bei 37 °C und 5 % CO2 für 48 h inkubiert, nachdem sie 24 h lang künstlichen Süßstoffen ausgesetzt waren. In der Zwischenzeit wurde eine einzelne Kolonie von E. coli und E. faecalis in 10 mL des jeweiligen Mediums, das mit den künstlichen Süßstoffen in Gegenwart oder Abwesenheit von Zinksulfat oder Vehikel (ddH2O) ergänzt wurde, beimpft und über Nacht bei 37 °C unter Schütteln bei 150 U/min inkubiert. Die Bakterien wurden dann zweimal mit 500 µL serum- und antibiotikafreiem EMEM-Medium durch Zentrifugieren bei 4000 U/min (2683× g) für 10 min bei 37 °C gewaschen (accuSpinTM 1R, Fisher Scientific, Thermo Electron Corporation LED GmbH, Osterode, Deutschland) und in EMEM ohne Antibiotika resuspendiert.
Caco-2-Zellmonolayer wurden zweimal mit 500 µL PBS gewaschen und dann wurde EMEM (490 µL; ohne Antibiotika) in jede Vertiefung gegeben. Die Gesamtzahl der adhärenten Caco-2-Zellen wurde durch eine Zellzählung ermittelt. Bakteriensuspension (10 µL) wurde auf die Caco-2-Zellen mit einer Multiplicity of Infection (MOI) von 1:300 für eine Infektionsinkubationszeit von 1 h gegeben. Nach der Infektionszeit wurden die Zellen zweimal mit 500 µL sterilem PBS gewaschen, die Caco-2-Zellen wurden mit 500 µL 0,5% Triton X-100 durch Auf- und Abpipettieren lysiert. Die Anzahl der lebensfähigen Bakterien wurde durch Ausbringen von seriellen Verdünnungen der Zellsuspension auf entsprechende Festmedien bestimmt, gefolgt von einer Inkubation über Nacht bei 37 °C und dem Zählen der koloniebildenden Einheiten. Die bakterielle Adhäsion wurde als Verhältnis der gesamten angehefteten Bakterien pro lebensfähige Caco-2-Zellen ausgedrückt (normalisiert auf 1). Jeder Assay wurde in dreifacher Ausführung mit einer aufeinanderfolgenden Passage von Caco-2-Zellen durchgeführt.
4.7. Invasions-Assay
Caco-2-Zellen wurden auf 24-Well-Gewebekulturplatten in einer Dichte von 7,5 × 104 Zellen/Vertiefung für 36 h ausgesät und anschließend für 24 h bei 37 °C in befeuchteter Umgebung mit 5 % CO2 den künstlichen Süßungsmitteln ausgesetzt. Der Zellmonolayer wurde zweimal mit 500 µL sterilem PBS gewaschen und 490 µL frisches EMEM-Medium ohne Antibiotika zugegeben. Die bakterielle Invasion von Caco-2-Zellen wurde mit dem zuvor beschriebenen Antibiotikaschutz-Assay 91Inaba, H.; Nomura, R.; Kato, Y.; Takeuchi, H.; Amano, A.; Asai, F.; Nakano, K.; Lamont, R.J.; Matsumoto-Nakano, M. Adhesion and invasion of gingival epithelial cells by Porphyromonas gulae. PLoS ONE 2019, 14, e0213309. [Google Scholar] [CrossRef] [PubMed] mit einigen Modifikationen gemessen. Die Bakterien wurden den künstlichen Süßungsmitteln ausgesetzt und zur Infektion vorbereitet, wie in Abschnitt 4.6 beschrieben. Die Anzahl der adhärenten Caco-2-Zellen, die einer bakteriellen Infektion ausgesetzt waren, wurde durch eine Zellzählung bestimmt. Der Caco-2-Zellmonolayer wurde mit Bakterien bei einer MOI von 1:300 für 1 h bei 37 °C infiziert. Der Monolayer wurde einmal mit 500 µL PBS gewaschen und frisches Zellkulturmedium (500 µL) zugegeben, das 100 µg/mL Gentamicin für E. coli und 100 µg/mL Gentamicin zusammen mit 50 µg/mL Ampicillin für E. faecalis enthielt, und für 30 min bei 37 °C inkubiert, um die extern anhaftenden Bakterien abzutöten 92Inaba, H.; Nomura, R.; Kato, Y.; Takeuchi, H.; Amano, A.; Asai, F.; Nakano, K.; Lamont, R.J.; Matsumoto-Nakano, M. Adhesion and invasion of gingival epithelial cells by Porphyromonas gulae. PLoS ONE 2019, 14, e0213309. [Google Scholar] [CrossRef] [PubMed]. Der Zellmonolayer wurde zweimal mit PBS gewaschen und anschließend mit 0,5 % Triton X-100 in PBS lysiert.
Die Anzahl der lebensfähigen koloniebildenden Einheiten wurde durch Verdünnen und Ausplattieren der Proben auf Festmedien und Inkubation über Nacht bei 37 °C bestimmt. Die Ergebnisse wurden als das Verhältnis der intrazellulären Bakterien im Vergleich zur Kontrolle ausgedrückt (normalisiert auf 1). Jeder Assay wurde in dreifacher Ausführung mit der sukzessiven Passage von Caco-2-Zellen durchgeführt.
4.8. Zytotoxizitäts-Assay
Die zytotoxische Wirkung von AS-vermittelten bakteriellen Metaboliten auf intestinale Epithelzellen wurde gemäß dem zuvor beschriebenen Protokoll 93Figueiredo, P.M.; Furumura, M.T.; Santos, A.M.; Sousa, A.C.; Kota, D.J.; Levy, C.E.; Yano, T. Cytotoxic activity of clinical Stenotrophomonas maltophilia. Lett. Appl. Microbiol. 2006, 43, 443–449. [Google Scholar] [CrossRef] [PubMed] mit Modifikationen durchgeführt, und die Zellviabilität wurde mit dem Cell Counting Kit-8 (CCK-8) gemäß den Richtlinien des Herstellers gemessen. Caco-2-Zellen wurden auf 96-Well-Platten (1 × 10⁴ Zellen/Well) gezüchtet und für 48 h bei 37 °C in befeuchteten Bedingungen mit 5 % CO2 inkubiert. Gleichzeitig wurde E. coli oder E. faecalis in 10 mL des jeweiligen Flüssigmediums, das mit 100 μM der künstlichen Süßstoffe mit oder ohne 100 μM/mL Zinksulfat oder Vehikel ergänzt wurde, für 24 h gezüchtet. Die Kulturen wurden bei 4000 U/min (2683× g) für 15 min bei 4 °C zentrifugiert und der Überstand wurde gesammelt und filtriert (0,22 µM-Membranen; Millipore, USA). 50 μL der löslichen bakteriellen Faktoren (Überstand) und 50 μL EMEM ohne Antibiotika wurden zu dem vorgewaschenen (mit 100 μL sterilem PBS) Caco-2 Zellmonolayer gegeben. Die Zellen wurden 24 h lang inkubiert und anschließend die Zellviabilität mit dem CCK-8-Reagenz gemessen, das als Absorption bei 450 nm mit einem Mikroplattenlesegerät (Tecan SunriseTM, Dänemark, Schweiz) bestimmt wurde.
4.9. Statistische Auswertung
Alle quantitativen Daten wurden als Excel-Dateien (Microsoft Office) erfasst. Der Durchschnitt der Triplikate für jede Studie wurde als n = 1 mit n = 5-6 für alle Studien außer dem Hämolyse-Assay analysiert. Die Daten wurden mit GraphPad Prism 7 analysiert. Die statistische Analyse wurde entweder mit einer einseitigen ANOVA oder einer zweiseitigen ANOVA durchgeführt, gegebenenfalls mit einem Tukey-Post-hoc-Test für multiple Vergleiche. Signifikanz war erreicht, wenn p < 0,05.
Autoren-Beiträge
A.S.: Konzeptualisierung, Methodik, Validierung, formale Analyse, Untersuchung, Datenkuratierung, Erstellung des ursprünglichen Entwurfs, schriftliche Überprüfung und Redaktion. H.C.: Konzeptualisierung, Ressourcen, formale Analyse, Verfassen des Originalentwurfs, Korrekturlesen und Lektorat, Supervision, Projektverwaltung. Alle Autoren haben die veröffentlichte Version des Manuskripts gelesen und sind damit einverstanden.
Finanzierung
Diese Forschung erhielt keine externe Finanzierung. Das APC wurde vom Anglia Ruskin University Open Access Fund finanziert.
Stellungnahme des Institutional Review Board
Es sind keine Tier- oder Humanstudien enthalten, jedoch wurde eine ethische Genehmigung für In-vitro-Studien vom Ethikausschuss der School of Life Sciences, Anglia Ruskin University, vom 23. März 2016 bis zum 1. Juni 2019 eingeholt.
Erklärung zur informierten Zustimmung
Nicht zutreffend.
Erklärung zur Datenverfügbarkeit
Datenfreigabe nicht anwendbar.
Interessenkonflikte
Die Autoren deklarieren keine Interessenkonflikte.
Copyright
© 2021 durch die Autoren. Lizenznehmer MDPI, Basel, Schweiz. Dieser Artikel ist ein Open-Access-Artikel, der unter den Bedingungen der Creative Commons Attribution (CC BY 4.0) Lizenz verbreitet wird (https://creativecommons.org/licenses/by/4.0/).
Info-Krümel
Die Bewertungs-Funktion ist derzeit deaktiviert!
Materialien zur Verfügung gestellt durch: Journals IJMS Volume 22 Issue 10 10.3390/ijms22105228
Inhalt wurde von w3punkt.de dem Original getreu ins Deutsche übersetzt und in Kernaussagen hervorgehoben.
JOURNAL-REFERENZ
https://www.mdpi.com/1422-0067/22/10/5228/htm#
TOOLTIPS
Die Texte in eventuell vorhandenen Tooltip-Fenstern wurden von der Redaktion des w3punkt.de bereitgestellt, sie entstammen in der Hauptsache den englisch- und deutschsprachigen Wikipedias.
Quellen und Tiefen
- 1Biomedical Research Group, School of Life Sciences, East Road, Cambridge CB1 1PT, UK
- 2School of Life Sciences, Anglia Ruskin University, Cambridge CB1 1PT, UK – Autor, an den die Korrespondenz gerichtet werden sollte.
- 3Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef]
- 4Wekerle, H. Brain autoimmunity and intestinal microbiota: 100 trillion game changers. Trends Immunol. 2017, 38, 483–497. [Google Scholar] [CrossRef]
- 5Li, J.; Jia, H.; Cai, X.; Zhong, H.; Feng, Q.; Sunagawa, S.; Arumugam, M.; Kultima, J.R.; Prifti, E.; Nielsen, T.; et al. An integrated catalog of reference genes in the human gut microbiome. Nat. Biotechnol. 2014, 32, 834–841. [Google Scholar] [CrossRef]
- 6Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed]
- 7Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, S.; Srinivasan, S.; Ley, R.E.; Gewirtz, A.T. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 2015, 519, 92–96. [Google Scholar] [CrossRef] [PubMed]
- 8Petersson, J.; Schreiber, O.; Hansson, G.C.; Gendler, S.J.; Velcich, A.; Lundberg, J.O.; Roos, S.; Holm, L.; Philipson, M. Importance and regulation of the colonic mucus barrier in a mouse model of colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, 2. [Google Scholar] [CrossRef] [PubMed]
- 9Vaishnava, S.; Behrendt, C.L.; Ismail, A.S.; Eckmann, L.; Hooper, L.V. Paneth cells directly sense gut commensals and maintain homeostasis at the intestinal host-microbial interface. Proc. Natl. Acad. Sci. USA 2008, 10552, 20858–20863. [Google Scholar] [CrossRef] [PubMed]
- 10Kelly, C.J.; Zheng, L.; Campbell, E.L.; Saeedi, B.; Scholz, C.C.; Bayless, A.J.; Wilson, K.E.; Glover, L.E.; Kominsky, D.J.; Magnuson, A.; et al. Crosstalk between microbiota-derived short-chain fatty acids and intestinal epithelial HIF augments tissue barrier function. Cell Host Microbe 2015, 175, 662–671. [Google Scholar] [CrossRef] [PubMed]
- 11Iraporda, C.; Errea, A.; Romanin, D.E.; Cayet, D.; Pereyra, E.; Pignataro, O.; Sirard, J.C.; Garrote, G.L.; Abraham, A.G.; Rumbo, M. Lactate and short chain fatty acids produced by microbial fermentation downregulate proinflammatory responses in intestinal epithelial cells and myeloid cells. Immunobiology 2015, 22010, 1161–1169. [Google Scholar] [CrossRef]
- 12de la Cuesta-Zuluaga, J.; Mueller, N.T.; Alvarez-Quintero, R.; Velasquez-Meija, E.P.; Sierra, J.A.; Corrales-Agudelo, V.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Higher fecal short-chain fatty acid levels are associated with gut microbiome dysbiosis, obesity, hypertension and cardiometabolic disease risk factors. Nutrients 2018, 11, 51. [Google Scholar] [CrossRef]
- 13LeBlanc, J.G.; Milani, C.; de Giori, G.S.; Sesma, F.; van Sinderen, D.; Ventura, M. Bacteria as vitamin suppliers to their host: A gut microbiota perspective. Curr. Opin. Biotechnol. 2013, 24, 160–168. [Google Scholar] [CrossRef]
- 14Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed]
- 15David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- 16De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed]
- 17Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed]
- 18Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, S.; Srinivasan, S.; Ley, R.E.; Gewirtz, A.T. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 2015, 519, 92–96. [Google Scholar] [CrossRef] [PubMed]
- 19Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]
- 20Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, S.; Srinivasan, S.; Ley, R.E.; Gewirtz, A.T. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 2015, 519, 92–96. [Google Scholar] [CrossRef] [PubMed]
- 21Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, S.; Srinivasan, S.; Ley, R.E.; Gewirtz, A.T. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 2015, 519, 92–96. [Google Scholar] [CrossRef] [PubMed]
- 22Gardner, C.; Wylie-Rosett, J.; Gidding, S.S.; Sen, L.M.; Johnson, R.K.; Reader, D.; Lichtenstein, A.H.; Physical Activity and Metabolism American Heart Association Nutrition Committee of the Council on Nutrition and American Diabetes Association. Nonnutritive sweeteners: Current use and health perspectives: A scientific statement from the american heart association and the american diabetes association. Diabetes Care 2012, 35, 1798–1808. [Google Scholar] [CrossRef]
- 23Gardner, C.; Wylie-Rosett, J.; Gidding, S.S.; Sen, L.M.; Johnson, R.K.; Reader, D.; Lichtenstein, A.H.; Physical Activity and Metabolism American Heart Association Nutrition Committee of the Council on Nutrition and American Diabetes Association. Nonnutritive sweeteners: Current use and health perspectives: A scientific statement from the american heart association and the american diabetes association. Diabetes Care 2012, 35, 1798–1808. [Google Scholar] [CrossRef]
- 24Blackburn, G.L.; Kanders, B.S.; Lavin, P.T.; Keller, S.D.; Whatley, J. The effect of aspartame as part of a multidisciplinary weight-control program on short- and long-term control of body weight. Am. J. Clin. Nutr. 1997, 65, 409–418. [Google Scholar] [CrossRef]
- 25Suez, J.; Korem, T.; Zilberman-Schapira, G.; Segal, E.; Elinav, E. Non-caloric artificial sweeteners and the microbiome: Findings and challenges. Gut Microbes 2015, 6, 149–155. [Google Scholar] [CrossRef]
- 26Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]
- 27De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed]
- 28Suez, J.; Korem, T.; Zilberman-Schapira, G.; Segal, E.; Elinav, E. Non-caloric artificial sweeteners and the microbiome: Findings and challenges. Gut Microbes 2015, 6, 149–155. [Google Scholar] [CrossRef]
- 29Frankenfeld, C.L.; Sikaroodi, M.; Lamb, E.; Shoemaker, S.; Gillevet, P.M. High-intensity sweetener consumption and gut microbiome content and predicted gene function in a cross-sectional study of adults in the united states. Ann. Epidemiol. 2015, 25, 736–742. [Google Scholar] [CrossRef]
- 30Bian, X.; Chi, L.; Gao, B.; Tu, P.; Ru, H.; Lu, K. Gut microbiome response to sucralose and its potential role in inducing liver inflammation in mice. Front. Physiol. 2017, 8, 487. [Google Scholar] [CrossRef]
- 31Bian, X.; Tu, P.; Chi, L.; Gao, B.; Ru, H.; Lu, K. Saccharin induced liver inflammation in mice by altering the gut microbiota and its metabolic functions. Food Chem. Toxicol. 2017, 107, 530–539. [Google Scholar] [CrossRef] [PubMed]
- 32Shil, A.; Olusanya, O.; Ghufoor, Z.; Forson, B.; Marks, J.; Chichger, H. Artificial sweeteners disrupt tight junctions and barrier function in the intestinal epithelium through activation of the sweet taste receptor, T1R3. Nutrients 2020, 12, 1862. [Google Scholar] [CrossRef]
- 33Mohamed, J.A.; Huang, D.B. Biofilm formation by enterococci. J. Med. Microbiol. 2007, 56, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- 34Tenaillon, O.; Skurnik, D.; Picard, B.; Denamur, E. The population genetics of commensal Escherichia coli. Nat. Rev. Microbiol. 2010, 8, 207–217. [Google Scholar] [CrossRef] [PubMed]
- 35Jimenez, E.; Marin, M.L.; Martin, R.; Odriozola, J.M.; Olivares, M.; Xaus, J.; Fernandez, L.; Rodriguez, J.M. Is meconium from healthy newborns actually sterile? Res. Microbiol. 2008, 159, 187–193. [Google Scholar] [CrossRef]
- 36Shil, A.; Olusanya, O.; Ghufoor, Z.; Forson, B.; Marks, J.; Chichger, H. Artificial sweeteners disrupt tight junctions and barrier function in the intestinal epithelium through activation of the sweet taste receptor, T1R3. Nutrients 2020, 12, 1862. [Google Scholar] [CrossRef]
- 37Mace, O.J.; Affleck, J.; Patel, N.; Kellett, G.L. Sweet taste receptors in rat small intestine stimulate glucose absorption through apical GLUT2. J. Physiol. 2007, 582, 379–392. [Google Scholar] [CrossRef]
- 38Keast, R.S.; Canty, T.M.; Breslin, P.A. Oral zinc sulfate solutions inhibit sweet taste perception. Chem. Senses 2004, 29, 513–521. [Google Scholar] [CrossRef]
- 39Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]
- 40Frankenfeld, C.L.; Sikaroodi, M.; Lamb, E.; Shoemaker, S.; Gillevet, P.M. High-intensity sweetener consumption and gut microbiome content and predicted gene function in a cross-sectional study of adults in the united states. Ann. Epidemiol. 2015, 25, 736–742. [Google Scholar] [CrossRef]
- 41Suez, J.; Korem, T.; Zilberman-Schapira, G.; Segal, E.; Elinav, E. Non-caloric artificial sweeteners and the microbiome: Findings and challenges. Gut Microbes 2015, 6, 149–155. [Google Scholar] [CrossRef]
- 42Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]
- 43Shil, A.; Olusanya, O.; Ghufoor, Z.; Forson, B.; Marks, J.; Chichger, H. Artificial sweeteners disrupt tight junctions and barrier function in the intestinal epithelium through activation of the sweet taste receptor, T1R3. Nutrients 2020, 12, 1862. [Google Scholar] [CrossRef]
- 44Wang, Q.P.; Browman, D.; Herzog, H.; Neely, G.G. Non-nutritive sweeteners possess a bacteriostatic effect and alter gut microbiota in mice. PLoS ONE 2018, 13, e0199080. [Google Scholar] [CrossRef]
- 45Gardner, C.; Wylie-Rosett, J.; Gidding, S.S.; Sen, L.M.; Johnson, R.K.; Reader, D.; Lichtenstein, A.H.; Physical Activity and Metabolism American Heart Association Nutrition Committee of the Council on Nutrition and American Diabetes Association. Nonnutritive sweeteners: Current use and health perspectives: A scientific statement from the american heart association and the american diabetes association. Diabetes Care 2012, 35, 1798–1808. [Google Scholar] [CrossRef]
- 46Ceri, H.; Olson, M.; Morck, D.; Storey, D.; Read, R.; Olson, B. The MBEC assay system: Multiple equivalent biofilms for antibiotic and biocide susceptibility testing. Methods Enzymol. 2001, 337, 377–385. [Google Scholar] [CrossRef]
- 47Kong, K.F.; Vuong, C.; Otto, M. Staphylococcus quorum sensing in biofilm formation and infection. Int. J. Med. Microbiol. 2006, 296, 133–139. [Google Scholar] [CrossRef] [PubMed]
- 48Wagner, V.E.; Li, L.L.; Isabella, V.M.; Iglewski, B.H. Analysis of the hierarchy of quorum-sensing regulation in Pseudomonas aeruginosa. Anal. Bioanal. Chem. 2007, 387, 469–479. [Google Scholar] [CrossRef] [PubMed]
- 49Hyland, R.M.; Sun, J.; Griener, T.P.; Mulvey, G.L.; Klassen, J.S.; Donnerberg, M.S.; Armstrong, G.D. The bundlin pilin protein of enteropathogenic Escherichia coli is an N-acetyllactosamine-specific lectin. Cell Microbiol. 2008, 10, 177–187. [Google Scholar] [CrossRef] [PubMed]
- 50Oscarsson, J.; Mizunoe, Y.; Li, L.; Lai, X.H.; Wieslander, A.; Uhlin, B.E. Molecular analysis of the cytolytic protein ClyA (SheA) from Escherichia coli. Mol. Microbiol. 1999, 32, 1226–1238. [Google Scholar] [CrossRef] [PubMed]
- 51Saldana, Z.; Erdem, A.L.; Schuller, S.; Okeke, I.N.; Lucas, M.; Sivananthan, A.; Philips, A.D.; Kaper, J.B.; Puente, J.L.; Giron, J.A. The Escherichia coli common pilus and the bundle-forming pilus act in concert during the formation of localized adherence by enteropathogenic E. coli. J. Bacteriol. 2009, 191, 3451–3461. [Google Scholar] [CrossRef]
- 52Huycke, M.M.; Joyce, W.A.; Gilmore, M.S. Enterococcus faecalis cytolysin without effect on the intestinal growth of susceptible enterococci in mice. J. Infect. Dis. 1995, 172, 273–276. [Google Scholar] [CrossRef]
- 53Theilacker, C.; Sanchez-Carballo, P.; Toma, I.; Fabretti, F.; Sava, I.; Kropec, A.; Holst, O.; Huebner, J. Glycolipids are involved in biofilm accumulation and prolonged bacteraemia in Enterococcus faecalis. Mol. Microbiol. 2009, 71, 1055–1069. [Google Scholar] [CrossRef]
- 54Wells, C.L.; Moore, E.A.; Hoag, J.A.; Hirt, H.; Dunny, G.M.; Erlandsen, S.L. Inducible expression of Enterococcus faecalis aggregation substance surface protein facilitates bacterial internalization by cultured enterocytes. Infect. Immun. 2000, 68, 7190–7194. [Google Scholar] [CrossRef]
- 55Sartingen, S.; Rozdzinski, E.; Muscholl-Silberhorn, A.; Marre, R. Aggregation substance increases adherence and internalization, but not translocation, of Enterococcus faecalis through different intestinal epithelial cells in vitro. Infect. Immun. 2000, 68, 6044–6047. [Google Scholar] [CrossRef]
- 56Ludwig, A.; Bauer, S.; Benz, R.; Bergmann, B.; Goebel, W. Analysis of the SlyA-controlled expression, subcellular localization and pore-forming activity of a 34 kDa haemolysin (ClyA) from Escherichia coli K-12. Mol. Microbiol. 1999, 31, 557–567. [Google Scholar] [CrossRef]
- 57Sheikh, J.; Hicks, S.; Dall’Agnol, M.; Phillips, A.D.; Nataro, J.P. Roles for fis and YafK in biofilm formation by enteroaggregative Escherichia coli. Mol. Microbiol. 2001, 41, 983–997. [Google Scholar] [CrossRef]
- 58Heikens, E.; Bonten, M.J.; Willems, R.J. Enterococcal surface protein esp is important for biofilm formation of Enterococcus faecium E1162. J. Bacteriol. 2007, 189, 8233–8240. [Google Scholar] [CrossRef]
- 59Srivastava, M.; Mallard, C.; Barke, T.; Hancock, L.E.; Self, W.T. A selenium-dependent xanthine dehydrogenase triggers biofilm proliferation in Enterococcus faecalis through oxidant production. J. Bacteriol. 2011, 193, 1643–1652. [Google Scholar] [CrossRef] [PubMed]
- 60Shankar, N.; Baghdayan, A.S.; Gilmore, M.S. Modulation of virulence within a pathogenicity island in vancomycin-resistant Enterococcus faecalis. Nature 2002, 417, 746–750. [Google Scholar] [CrossRef]
- 61Shankar, N.; Baghdayan, A.S.; Gilmore, M.S. Modulation of virulence within a pathogenicity island in vancomycin-resistant Enterococcus faecalis. Nature 2002, 417, 746–750. [Google Scholar] [CrossRef]
- 62Giaffer, M.H.; Holdsworth, C.D.; Duerden, B.I. Virulence properties of Escherichia coli strains isolated from patients with inflammatory bowel disease. Gut 1992, 33, 646–650. [Google Scholar] [CrossRef]
- 63Ike, Y.; Hashimoto, H.; Clewell, D.B. High incidence of hemolysin production by Enterococcus (streptococcus) faecalis strains associated with human parenteral infections. J. Clin. Microbiol. 1987, 25, 1524–1528. [Google Scholar] [CrossRef] [PubMed]
- 64Mirsepasi-Lauridsen, H.C.; Du, Z.; Struve, C.; Charbon, G.; Karczewski, J.; Krogfelt, K.A.; Petersen, A.M.; Wells, J.M. Secretion of alpha-hemolysin by Escherichia coli disrupts tight junctions in ulcerative colitis patients. Clin. Transl. Gastroenterol. 2016, 7, e149. [Google Scholar] [CrossRef] [PubMed]
- 65Nelson, G.; Hoon, M.A.; Chandrashekar, J.; Zhang, Y.; Ryba, N.J.; Zuker, C.S. Mammalian sweet taste receptors. Cell 2001, 106, 381–390. [Google Scholar] [CrossRef]
- 66Keast, R.S.; Canty, T.M.; Breslin, P.A. Oral zinc sulfate solutions inhibit sweet taste perception. Chem. Senses 2004, 29, 513–521. [Google Scholar] [CrossRef]
- 67Wee, M.; Tan, V.; Forde, C. A comparison of psychophysical dose-response behaviour across 16 sweeteners. Nutrients 2018, 10, 1632. [Google Scholar] [CrossRef] [PubMed]
- 68Nijland, R.; Burgess, J.G. Bacterial olfaction. Biotechnol. J. 2010, 5, 974–977. [Google Scholar] [CrossRef]
- 69Bandyopadhyay, A.; Ghoshal, S.; Mukherjee, A. Genotoxicity testing of low-calorie sweeteners: Aspartame, acesulfame-K., and saccharin. Drug Chem. Toxicol. 2008, 31, 447–457. [Google Scholar] [CrossRef] [PubMed]
- 70Qu, Y.; Li, R.; Jiang, M.; Wang, X. Sucralose increases antimicrobial resistance and stimulates recovery of Escherichia coli mutants. Curr. Microbiol. 2017, 74, 885–888. [Google Scholar] [CrossRef] [PubMed]
- 71Rencuzogullari, E.; Tuylu, B.A.; Topaktas, M.; Ila, H.B.; Kayraldiz, A.; Arslan, M.; Diler, S.B. Genotoxicity of aspartame. Drug Chem. Toxicol. 2004, 27, 257–268. [Google Scholar] [CrossRef]
- 72Yu, Z.; Wang, Y.; Lu, J.; Bond, P.L.; Guo, J. Nonnutritive sweeteners can promote the dissemination of antibiotic resistance through conjugative gene transfer. ISME J. 2021. [Google Scholar] [CrossRef] [PubMed]
- 73Frey, R.L.; He, L.; Cui, Y.; Decho, A.W.; Kawaguchi, T.; Fergusonm, P.L.; Ferry, J.L. Reaction of N-acylhomoserine lactones with hydroxyl radicals: Rates, products, and effects on signaling activity. Environ. Sci. Technol. 2010, 44, 7465–7469. [Google Scholar] [CrossRef]
- 74Rothfork, J.M.; Timmins, G.S.; Harris, M.N.; Chen, X.; Lusis, A.J.; Otto, M.; Cheung, A.L.; Gresham, D. Inactivation of a bacterial virulence pheromone by phagocyte-derived oxidants: New role for the NADPH oxidase in host defense. Proc. Natl. Acad. Sci. USA 2004, 101, 13867–13872. [Google Scholar] [CrossRef]
- 75Ghoshal, U.C.; Ghoshal, U.; Jain, M.; Kumar, A.; Aggarwal, R.; Misra, A.; Ayyagari, A.; Naik, S.R. Strongyloides stercoralis infestation associated with septicemia due to intestinal transmural migration of bacteria. J. Gastroenterol. Hepatol. 2002, 17, 1331–1333. [Google Scholar] [CrossRef]
- 76Wells, C.L.; Jechorek, R.P.; Erlandsen, S.L. Evidence for the translocation of Enterococcus faecalis across the mouse intestinal tract. J. Infect. Dis. 1990, 162, 82–90. [Google Scholar] [CrossRef]
- 77Wells, C.L.; Erlandsen, S.L. Localization of translocating Escherichia coli, proteus mirabilis, and Enterococcus faecalis within cecal and colonic tissues of monoassociated mice. Infect. Immun. 1991, 59, 4693–4697. [Google Scholar] [CrossRef]
- 78Shil, A.; Olusanya, O.; Ghufoor, Z.; Forson, B.; Marks, J.; Chichger, H. Artificial sweeteners disrupt tight junctions and barrier function in the intestinal epithelium through activation of the sweet taste receptor, T1R3. Nutrients 2020, 12, 1862. [Google Scholar] [CrossRef]
- 79Santos, P.S.; Caria, C.R.P.; Gotardo, E.M.F.; Ribeiro, M.L.; Pedrazzoli, J.; Gambero, A. Artificial sweetener saccharin disrupts intestinal epithelial cells’ barrier function in vitro. Food Funct. 2018, 9, 3815–3822. [Google Scholar] [CrossRef]
- 80Yang, Y.Y.; Liu, W.R.; Liu, Y.S.; Zhao, J.L.; Zhang, Q.Q.; Zhang, M.; Zhang, J.N.; Jiang, Y.X.; Zhang, L.J.; Ying, G.G. Suitability of pharmaceuticals and personal care products (PPCPs) and artificial sweeteners (ASs) as wastewater indicators in the pearl river delta, south china. Sci. Total Environ. 2017, 590–591, 611–619. [Google Scholar] [CrossRef]
- 81Tran, N.H.; Hu, J.; Li, J.; Ong, S.L. Suitability of artificial sweeteners as indicators of raw wastewater contamination in surface water and groundwater. Water Res. 2014, 48, 443–456. [Google Scholar] [CrossRef]
- 82Tran, N.H.; Gan, J.; Nguyen, V.T.; Chen, H.; You, L.; Duarah, A.; Zhang, L.; Gin, K.Y. Sorption and biodegradation of artificial sweeteners in activated sludge processes. Bioresour. Technol. 2015, 197, 329–338. [Google Scholar] [CrossRef] [PubMed]
- 83Qu, Y.; Li, R.; Jiang, M.; Wang, X. Sucralose increases antimicrobial resistance and stimulates recovery of Escherichia coli mutants. Curr. Microbiol. 2017, 74, 885–888. [Google Scholar] [CrossRef] [PubMed]
- 84Yu, Z.; Wang, Y.; Lu, J.; Bond, P.L.; Guo, J. Nonnutritive sweeteners can promote the dissemination of antibiotic resistance through conjugative gene transfer. ISME J. 2021. [Google Scholar] [CrossRef] [PubMed]
- 85Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- 86Lleo, M.; Bonato, B.; Tafi, M.C.; Caburlotto, G.; Benedetti, D.; Canepari, P. Adhesion to medical device materials and biofilm formation capability of some species of enterococci in different physiological states. FEMS Microbiol. Lett. 2007, 274, 232–237. [Google Scholar] [CrossRef] [PubMed]
- 87Mack, D.; Rohde, H.; Harris, L.G.; Davies, A.P.; Horstkotte, M.A.; Knobloch, J.K. Biofilm formation in medical device-related infection. Int. J. Artif. Organs. 2006, 29, 343–359. [Google Scholar] [CrossRef] [PubMed]
- 88Braxton, E.E.; Ehrlich, G.D.; Hall-Stoodley, L.; Stoodley, P.; Veeh, R.; Fux, C.; Hu, F.Z.; Quigley, M.; Post, J.C. Role of biofilms in neurosurgical device-related infections. Neurosurg. Rev. 2005, 28, 249–255. [Google Scholar] [CrossRef]
- 89Wiseman, G.M. The hemolysins of Staphylococcus aureus. Bacteriol. Rev. 1975, 39, 317–344. [Google Scholar] [CrossRef] [PubMed]
- 90Darfeuille-Michaud, A.; Aubel, D.; Chauviere, G.; Rich, C.; Bourges, M.; Servin, A.; Joly, B. Adhesion of enterotoxigenic Escherichia coli to the human colon carcinoma cell line caco-2 in culture. Infect. Immun. 1990, 58, 893–902. [Google Scholar] [CrossRef]
- 91Inaba, H.; Nomura, R.; Kato, Y.; Takeuchi, H.; Amano, A.; Asai, F.; Nakano, K.; Lamont, R.J.; Matsumoto-Nakano, M. Adhesion and invasion of gingival epithelial cells by Porphyromonas gulae. PLoS ONE 2019, 14, e0213309. [Google Scholar] [CrossRef] [PubMed]
- 92Inaba, H.; Nomura, R.; Kato, Y.; Takeuchi, H.; Amano, A.; Asai, F.; Nakano, K.; Lamont, R.J.; Matsumoto-Nakano, M. Adhesion and invasion of gingival epithelial cells by Porphyromonas gulae. PLoS ONE 2019, 14, e0213309. [Google Scholar] [CrossRef] [PubMed]
- 93Figueiredo, P.M.; Furumura, M.T.; Santos, A.M.; Sousa, A.C.; Kota, D.J.; Levy, C.E.; Yano, T. Cytotoxic activity of clinical Stenotrophomonas maltophilia. Lett. Appl. Microbiol. 2006, 43, 443–449. [Google Scholar] [CrossRef] [PubMed]
Das Titelbild entstammen der Studie.
© 2021 durch die Autoren. Lizenznehmer MDPI, Basel, Schweiz. Dieser Artikel ist ein Open-Access-Artikel, der unter den Bedingungen der Creative Commons Attribution (CC BY 4.0) Lizenz verbreitet wird (https://creativecommons.org/licenses/by/4.0/).
Quellen und Tiefen
- 1Biomedical Research Group, School of Life Sciences, East Road, Cambridge CB1 1PT, UK
- 2School of Life Sciences, Anglia Ruskin University, Cambridge CB1 1PT, UK – Autor, an den die Korrespondenz gerichtet werden sollte.
- 3Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef]
- 4Wekerle, H. Brain autoimmunity and intestinal microbiota: 100 trillion game changers. Trends Immunol. 2017, 38, 483–497. [Google Scholar] [CrossRef]
- 5Li, J.; Jia, H.; Cai, X.; Zhong, H.; Feng, Q.; Sunagawa, S.; Arumugam, M.; Kultima, J.R.; Prifti, E.; Nielsen, T.; et al. An integrated catalog of reference genes in the human gut microbiome. Nat. Biotechnol. 2014, 32, 834–841. [Google Scholar] [CrossRef]
- 6Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed]
- 7Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, S.; Srinivasan, S.; Ley, R.E.; Gewirtz, A.T. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 2015, 519, 92–96. [Google Scholar] [CrossRef] [PubMed]
- 8Petersson, J.; Schreiber, O.; Hansson, G.C.; Gendler, S.J.; Velcich, A.; Lundberg, J.O.; Roos, S.; Holm, L.; Philipson, M. Importance and regulation of the colonic mucus barrier in a mouse model of colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, 2. [Google Scholar] [CrossRef] [PubMed]
- 9Vaishnava, S.; Behrendt, C.L.; Ismail, A.S.; Eckmann, L.; Hooper, L.V. Paneth cells directly sense gut commensals and maintain homeostasis at the intestinal host-microbial interface. Proc. Natl. Acad. Sci. USA 2008, 10552, 20858–20863. [Google Scholar] [CrossRef] [PubMed]
- 10Kelly, C.J.; Zheng, L.; Campbell, E.L.; Saeedi, B.; Scholz, C.C.; Bayless, A.J.; Wilson, K.E.; Glover, L.E.; Kominsky, D.J.; Magnuson, A.; et al. Crosstalk between microbiota-derived short-chain fatty acids and intestinal epithelial HIF augments tissue barrier function. Cell Host Microbe 2015, 175, 662–671. [Google Scholar] [CrossRef] [PubMed]
- 11Iraporda, C.; Errea, A.; Romanin, D.E.; Cayet, D.; Pereyra, E.; Pignataro, O.; Sirard, J.C.; Garrote, G.L.; Abraham, A.G.; Rumbo, M. Lactate and short chain fatty acids produced by microbial fermentation downregulate proinflammatory responses in intestinal epithelial cells and myeloid cells. Immunobiology 2015, 22010, 1161–1169. [Google Scholar] [CrossRef]
- 12de la Cuesta-Zuluaga, J.; Mueller, N.T.; Alvarez-Quintero, R.; Velasquez-Meija, E.P.; Sierra, J.A.; Corrales-Agudelo, V.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Higher fecal short-chain fatty acid levels are associated with gut microbiome dysbiosis, obesity, hypertension and cardiometabolic disease risk factors. Nutrients 2018, 11, 51. [Google Scholar] [CrossRef]
- 13LeBlanc, J.G.; Milani, C.; de Giori, G.S.; Sesma, F.; van Sinderen, D.; Ventura, M. Bacteria as vitamin suppliers to their host: A gut microbiota perspective. Curr. Opin. Biotechnol. 2013, 24, 160–168. [Google Scholar] [CrossRef]
- 14Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed]
- 15David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- 16De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed]
- 17Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed]
- 18Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, S.; Srinivasan, S.; Ley, R.E.; Gewirtz, A.T. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 2015, 519, 92–96. [Google Scholar] [CrossRef] [PubMed]
- 19Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]
- 20Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, S.; Srinivasan, S.; Ley, R.E.; Gewirtz, A.T. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 2015, 519, 92–96. [Google Scholar] [CrossRef] [PubMed]
- 21Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, S.; Srinivasan, S.; Ley, R.E.; Gewirtz, A.T. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 2015, 519, 92–96. [Google Scholar] [CrossRef] [PubMed]
- 22Gardner, C.; Wylie-Rosett, J.; Gidding, S.S.; Sen, L.M.; Johnson, R.K.; Reader, D.; Lichtenstein, A.H.; Physical Activity and Metabolism American Heart Association Nutrition Committee of the Council on Nutrition and American Diabetes Association. Nonnutritive sweeteners: Current use and health perspectives: A scientific statement from the american heart association and the american diabetes association. Diabetes Care 2012, 35, 1798–1808. [Google Scholar] [CrossRef]
- 23Gardner, C.; Wylie-Rosett, J.; Gidding, S.S.; Sen, L.M.; Johnson, R.K.; Reader, D.; Lichtenstein, A.H.; Physical Activity and Metabolism American Heart Association Nutrition Committee of the Council on Nutrition and American Diabetes Association. Nonnutritive sweeteners: Current use and health perspectives: A scientific statement from the american heart association and the american diabetes association. Diabetes Care 2012, 35, 1798–1808. [Google Scholar] [CrossRef]
- 24Blackburn, G.L.; Kanders, B.S.; Lavin, P.T.; Keller, S.D.; Whatley, J. The effect of aspartame as part of a multidisciplinary weight-control program on short- and long-term control of body weight. Am. J. Clin. Nutr. 1997, 65, 409–418. [Google Scholar] [CrossRef]
- 25Suez, J.; Korem, T.; Zilberman-Schapira, G.; Segal, E.; Elinav, E. Non-caloric artificial sweeteners and the microbiome: Findings and challenges. Gut Microbes 2015, 6, 149–155. [Google Scholar] [CrossRef]
- 26Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]
- 27De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed]
- 28Suez, J.; Korem, T.; Zilberman-Schapira, G.; Segal, E.; Elinav, E. Non-caloric artificial sweeteners and the microbiome: Findings and challenges. Gut Microbes 2015, 6, 149–155. [Google Scholar] [CrossRef]
- 29Frankenfeld, C.L.; Sikaroodi, M.; Lamb, E.; Shoemaker, S.; Gillevet, P.M. High-intensity sweetener consumption and gut microbiome content and predicted gene function in a cross-sectional study of adults in the united states. Ann. Epidemiol. 2015, 25, 736–742. [Google Scholar] [CrossRef]
- 30Bian, X.; Chi, L.; Gao, B.; Tu, P.; Ru, H.; Lu, K. Gut microbiome response to sucralose and its potential role in inducing liver inflammation in mice. Front. Physiol. 2017, 8, 487. [Google Scholar] [CrossRef]
- 31Bian, X.; Tu, P.; Chi, L.; Gao, B.; Ru, H.; Lu, K. Saccharin induced liver inflammation in mice by altering the gut microbiota and its metabolic functions. Food Chem. Toxicol. 2017, 107, 530–539. [Google Scholar] [CrossRef] [PubMed]
- 32Shil, A.; Olusanya, O.; Ghufoor, Z.; Forson, B.; Marks, J.; Chichger, H. Artificial sweeteners disrupt tight junctions and barrier function in the intestinal epithelium through activation of the sweet taste receptor, T1R3. Nutrients 2020, 12, 1862. [Google Scholar] [CrossRef]
- 33Mohamed, J.A.; Huang, D.B. Biofilm formation by enterococci. J. Med. Microbiol. 2007, 56, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- 34Tenaillon, O.; Skurnik, D.; Picard, B.; Denamur, E. The population genetics of commensal Escherichia coli. Nat. Rev. Microbiol. 2010, 8, 207–217. [Google Scholar] [CrossRef] [PubMed]
- 35Jimenez, E.; Marin, M.L.; Martin, R.; Odriozola, J.M.; Olivares, M.; Xaus, J.; Fernandez, L.; Rodriguez, J.M. Is meconium from healthy newborns actually sterile? Res. Microbiol. 2008, 159, 187–193. [Google Scholar] [CrossRef]
- 36Shil, A.; Olusanya, O.; Ghufoor, Z.; Forson, B.; Marks, J.; Chichger, H. Artificial sweeteners disrupt tight junctions and barrier function in the intestinal epithelium through activation of the sweet taste receptor, T1R3. Nutrients 2020, 12, 1862. [Google Scholar] [CrossRef]
- 37Mace, O.J.; Affleck, J.; Patel, N.; Kellett, G.L. Sweet taste receptors in rat small intestine stimulate glucose absorption through apical GLUT2. J. Physiol. 2007, 582, 379–392. [Google Scholar] [CrossRef]
- 38Keast, R.S.; Canty, T.M.; Breslin, P.A. Oral zinc sulfate solutions inhibit sweet taste perception. Chem. Senses 2004, 29, 513–521. [Google Scholar] [CrossRef]
- 39Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]
- 40Frankenfeld, C.L.; Sikaroodi, M.; Lamb, E.; Shoemaker, S.; Gillevet, P.M. High-intensity sweetener consumption and gut microbiome content and predicted gene function in a cross-sectional study of adults in the united states. Ann. Epidemiol. 2015, 25, 736–742. [Google Scholar] [CrossRef]
- 41Suez, J.; Korem, T.; Zilberman-Schapira, G.; Segal, E.; Elinav, E. Non-caloric artificial sweeteners and the microbiome: Findings and challenges. Gut Microbes 2015, 6, 149–155. [Google Scholar] [CrossRef]
- 42Suez, J.; Korem, T.; Zeevi, D.; Zilberman-Schapira, G.; Thaiss, C.A.; Maza, O.; Israeli, D.; Zmora, N.; Gilad, S.; Weinberger, A.; et al. Artificial sweeteners induce glucose intolerance by altering the gut microbiota. Nature 2014, 514, 181–186. [Google Scholar] [CrossRef]
- 43Shil, A.; Olusanya, O.; Ghufoor, Z.; Forson, B.; Marks, J.; Chichger, H. Artificial sweeteners disrupt tight junctions and barrier function in the intestinal epithelium through activation of the sweet taste receptor, T1R3. Nutrients 2020, 12, 1862. [Google Scholar] [CrossRef]
- 44Wang, Q.P.; Browman, D.; Herzog, H.; Neely, G.G. Non-nutritive sweeteners possess a bacteriostatic effect and alter gut microbiota in mice. PLoS ONE 2018, 13, e0199080. [Google Scholar] [CrossRef]
- 45Gardner, C.; Wylie-Rosett, J.; Gidding, S.S.; Sen, L.M.; Johnson, R.K.; Reader, D.; Lichtenstein, A.H.; Physical Activity and Metabolism American Heart Association Nutrition Committee of the Council on Nutrition and American Diabetes Association. Nonnutritive sweeteners: Current use and health perspectives: A scientific statement from the american heart association and the american diabetes association. Diabetes Care 2012, 35, 1798–1808. [Google Scholar] [CrossRef]
- 46Ceri, H.; Olson, M.; Morck, D.; Storey, D.; Read, R.; Olson, B. The MBEC assay system: Multiple equivalent biofilms for antibiotic and biocide susceptibility testing. Methods Enzymol. 2001, 337, 377–385. [Google Scholar] [CrossRef]
- 47Kong, K.F.; Vuong, C.; Otto, M. Staphylococcus quorum sensing in biofilm formation and infection. Int. J. Med. Microbiol. 2006, 296, 133–139. [Google Scholar] [CrossRef] [PubMed]
- 48Wagner, V.E.; Li, L.L.; Isabella, V.M.; Iglewski, B.H. Analysis of the hierarchy of quorum-sensing regulation in Pseudomonas aeruginosa. Anal. Bioanal. Chem. 2007, 387, 469–479. [Google Scholar] [CrossRef] [PubMed]
- 49Hyland, R.M.; Sun, J.; Griener, T.P.; Mulvey, G.L.; Klassen, J.S.; Donnerberg, M.S.; Armstrong, G.D. The bundlin pilin protein of enteropathogenic Escherichia coli is an N-acetyllactosamine-specific lectin. Cell Microbiol. 2008, 10, 177–187. [Google Scholar] [CrossRef] [PubMed]
- 50Oscarsson, J.; Mizunoe, Y.; Li, L.; Lai, X.H.; Wieslander, A.; Uhlin, B.E. Molecular analysis of the cytolytic protein ClyA (SheA) from Escherichia coli. Mol. Microbiol. 1999, 32, 1226–1238. [Google Scholar] [CrossRef] [PubMed]
- 51Saldana, Z.; Erdem, A.L.; Schuller, S.; Okeke, I.N.; Lucas, M.; Sivananthan, A.; Philips, A.D.; Kaper, J.B.; Puente, J.L.; Giron, J.A. The Escherichia coli common pilus and the bundle-forming pilus act in concert during the formation of localized adherence by enteropathogenic E. coli. J. Bacteriol. 2009, 191, 3451–3461. [Google Scholar] [CrossRef]
- 52Huycke, M.M.; Joyce, W.A.; Gilmore, M.S. Enterococcus faecalis cytolysin without effect on the intestinal growth of susceptible enterococci in mice. J. Infect. Dis. 1995, 172, 273–276. [Google Scholar] [CrossRef]
- 53Theilacker, C.; Sanchez-Carballo, P.; Toma, I.; Fabretti, F.; Sava, I.; Kropec, A.; Holst, O.; Huebner, J. Glycolipids are involved in biofilm accumulation and prolonged bacteraemia in Enterococcus faecalis. Mol. Microbiol. 2009, 71, 1055–1069. [Google Scholar] [CrossRef]
- 54Wells, C.L.; Moore, E.A.; Hoag, J.A.; Hirt, H.; Dunny, G.M.; Erlandsen, S.L. Inducible expression of Enterococcus faecalis aggregation substance surface protein facilitates bacterial internalization by cultured enterocytes. Infect. Immun. 2000, 68, 7190–7194. [Google Scholar] [CrossRef]
- 55Sartingen, S.; Rozdzinski, E.; Muscholl-Silberhorn, A.; Marre, R. Aggregation substance increases adherence and internalization, but not translocation, of Enterococcus faecalis through different intestinal epithelial cells in vitro. Infect. Immun. 2000, 68, 6044–6047. [Google Scholar] [CrossRef]
- 56Ludwig, A.; Bauer, S.; Benz, R.; Bergmann, B.; Goebel, W. Analysis of the SlyA-controlled expression, subcellular localization and pore-forming activity of a 34 kDa haemolysin (ClyA) from Escherichia coli K-12. Mol. Microbiol. 1999, 31, 557–567. [Google Scholar] [CrossRef]
- 57Sheikh, J.; Hicks, S.; Dall’Agnol, M.; Phillips, A.D.; Nataro, J.P. Roles for fis and YafK in biofilm formation by enteroaggregative Escherichia coli. Mol. Microbiol. 2001, 41, 983–997. [Google Scholar] [CrossRef]
- 58Heikens, E.; Bonten, M.J.; Willems, R.J. Enterococcal surface protein esp is important for biofilm formation of Enterococcus faecium E1162. J. Bacteriol. 2007, 189, 8233–8240. [Google Scholar] [CrossRef]
- 59Srivastava, M.; Mallard, C.; Barke, T.; Hancock, L.E.; Self, W.T. A selenium-dependent xanthine dehydrogenase triggers biofilm proliferation in Enterococcus faecalis through oxidant production. J. Bacteriol. 2011, 193, 1643–1652. [Google Scholar] [CrossRef] [PubMed]
- 60Shankar, N.; Baghdayan, A.S.; Gilmore, M.S. Modulation of virulence within a pathogenicity island in vancomycin-resistant Enterococcus faecalis. Nature 2002, 417, 746–750. [Google Scholar] [CrossRef]
- 61Shankar, N.; Baghdayan, A.S.; Gilmore, M.S. Modulation of virulence within a pathogenicity island in vancomycin-resistant Enterococcus faecalis. Nature 2002, 417, 746–750. [Google Scholar] [CrossRef]
- 62Giaffer, M.H.; Holdsworth, C.D.; Duerden, B.I. Virulence properties of Escherichia coli strains isolated from patients with inflammatory bowel disease. Gut 1992, 33, 646–650. [Google Scholar] [CrossRef]
- 63Ike, Y.; Hashimoto, H.; Clewell, D.B. High incidence of hemolysin production by Enterococcus (streptococcus) faecalis strains associated with human parenteral infections. J. Clin. Microbiol. 1987, 25, 1524–1528. [Google Scholar] [CrossRef] [PubMed]
- 64Mirsepasi-Lauridsen, H.C.; Du, Z.; Struve, C.; Charbon, G.; Karczewski, J.; Krogfelt, K.A.; Petersen, A.M.; Wells, J.M. Secretion of alpha-hemolysin by Escherichia coli disrupts tight junctions in ulcerative colitis patients. Clin. Transl. Gastroenterol. 2016, 7, e149. [Google Scholar] [CrossRef] [PubMed]
- 65Nelson, G.; Hoon, M.A.; Chandrashekar, J.; Zhang, Y.; Ryba, N.J.; Zuker, C.S. Mammalian sweet taste receptors. Cell 2001, 106, 381–390. [Google Scholar] [CrossRef]
- 66Keast, R.S.; Canty, T.M.; Breslin, P.A. Oral zinc sulfate solutions inhibit sweet taste perception. Chem. Senses 2004, 29, 513–521. [Google Scholar] [CrossRef]
- 67Wee, M.; Tan, V.; Forde, C. A comparison of psychophysical dose-response behaviour across 16 sweeteners. Nutrients 2018, 10, 1632. [Google Scholar] [CrossRef] [PubMed]
- 68Nijland, R.; Burgess, J.G. Bacterial olfaction. Biotechnol. J. 2010, 5, 974–977. [Google Scholar] [CrossRef]
- 69Bandyopadhyay, A.; Ghoshal, S.; Mukherjee, A. Genotoxicity testing of low-calorie sweeteners: Aspartame, acesulfame-K., and saccharin. Drug Chem. Toxicol. 2008, 31, 447–457. [Google Scholar] [CrossRef] [PubMed]
- 70Qu, Y.; Li, R.; Jiang, M.; Wang, X. Sucralose increases antimicrobial resistance and stimulates recovery of Escherichia coli mutants. Curr. Microbiol. 2017, 74, 885–888. [Google Scholar] [CrossRef] [PubMed]
- 71Rencuzogullari, E.; Tuylu, B.A.; Topaktas, M.; Ila, H.B.; Kayraldiz, A.; Arslan, M.; Diler, S.B. Genotoxicity of aspartame. Drug Chem. Toxicol. 2004, 27, 257–268. [Google Scholar] [CrossRef]
- 72Yu, Z.; Wang, Y.; Lu, J.; Bond, P.L.; Guo, J. Nonnutritive sweeteners can promote the dissemination of antibiotic resistance through conjugative gene transfer. ISME J. 2021. [Google Scholar] [CrossRef] [PubMed]
- 73Frey, R.L.; He, L.; Cui, Y.; Decho, A.W.; Kawaguchi, T.; Fergusonm, P.L.; Ferry, J.L. Reaction of N-acylhomoserine lactones with hydroxyl radicals: Rates, products, and effects on signaling activity. Environ. Sci. Technol. 2010, 44, 7465–7469. [Google Scholar] [CrossRef]
- 74Rothfork, J.M.; Timmins, G.S.; Harris, M.N.; Chen, X.; Lusis, A.J.; Otto, M.; Cheung, A.L.; Gresham, D. Inactivation of a bacterial virulence pheromone by phagocyte-derived oxidants: New role for the NADPH oxidase in host defense. Proc. Natl. Acad. Sci. USA 2004, 101, 13867–13872. [Google Scholar] [CrossRef]
- 75Ghoshal, U.C.; Ghoshal, U.; Jain, M.; Kumar, A.; Aggarwal, R.; Misra, A.; Ayyagari, A.; Naik, S.R. Strongyloides stercoralis infestation associated with septicemia due to intestinal transmural migration of bacteria. J. Gastroenterol. Hepatol. 2002, 17, 1331–1333. [Google Scholar] [CrossRef]
- 76Wells, C.L.; Jechorek, R.P.; Erlandsen, S.L. Evidence for the translocation of Enterococcus faecalis across the mouse intestinal tract. J. Infect. Dis. 1990, 162, 82–90. [Google Scholar] [CrossRef]
- 77Wells, C.L.; Erlandsen, S.L. Localization of translocating Escherichia coli, proteus mirabilis, and Enterococcus faecalis within cecal and colonic tissues of monoassociated mice. Infect. Immun. 1991, 59, 4693–4697. [Google Scholar] [CrossRef]
- 78Shil, A.; Olusanya, O.; Ghufoor, Z.; Forson, B.; Marks, J.; Chichger, H. Artificial sweeteners disrupt tight junctions and barrier function in the intestinal epithelium through activation of the sweet taste receptor, T1R3. Nutrients 2020, 12, 1862. [Google Scholar] [CrossRef]
- 79Santos, P.S.; Caria, C.R.P.; Gotardo, E.M.F.; Ribeiro, M.L.; Pedrazzoli, J.; Gambero, A. Artificial sweetener saccharin disrupts intestinal epithelial cells’ barrier function in vitro. Food Funct. 2018, 9, 3815–3822. [Google Scholar] [CrossRef]
- 80Yang, Y.Y.; Liu, W.R.; Liu, Y.S.; Zhao, J.L.; Zhang, Q.Q.; Zhang, M.; Zhang, J.N.; Jiang, Y.X.; Zhang, L.J.; Ying, G.G. Suitability of pharmaceuticals and personal care products (PPCPs) and artificial sweeteners (ASs) as wastewater indicators in the pearl river delta, south china. Sci. Total Environ. 2017, 590–591, 611–619. [Google Scholar] [CrossRef]
- 81Tran, N.H.; Hu, J.; Li, J.; Ong, S.L. Suitability of artificial sweeteners as indicators of raw wastewater contamination in surface water and groundwater. Water Res. 2014, 48, 443–456. [Google Scholar] [CrossRef]
- 82Tran, N.H.; Gan, J.; Nguyen, V.T.; Chen, H.; You, L.; Duarah, A.; Zhang, L.; Gin, K.Y. Sorption and biodegradation of artificial sweeteners in activated sludge processes. Bioresour. Technol. 2015, 197, 329–338. [Google Scholar] [CrossRef] [PubMed]
- 83Qu, Y.; Li, R.; Jiang, M.; Wang, X. Sucralose increases antimicrobial resistance and stimulates recovery of Escherichia coli mutants. Curr. Microbiol. 2017, 74, 885–888. [Google Scholar] [CrossRef] [PubMed]
- 84Yu, Z.; Wang, Y.; Lu, J.; Bond, P.L.; Guo, J. Nonnutritive sweeteners can promote the dissemination of antibiotic resistance through conjugative gene transfer. ISME J. 2021. [Google Scholar] [CrossRef] [PubMed]
- 85Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- 86Lleo, M.; Bonato, B.; Tafi, M.C.; Caburlotto, G.; Benedetti, D.; Canepari, P. Adhesion to medical device materials and biofilm formation capability of some species of enterococci in different physiological states. FEMS Microbiol. Lett. 2007, 274, 232–237. [Google Scholar] [CrossRef] [PubMed]
- 87Mack, D.; Rohde, H.; Harris, L.G.; Davies, A.P.; Horstkotte, M.A.; Knobloch, J.K. Biofilm formation in medical device-related infection. Int. J. Artif. Organs. 2006, 29, 343–359. [Google Scholar] [CrossRef] [PubMed]
- 88Braxton, E.E.; Ehrlich, G.D.; Hall-Stoodley, L.; Stoodley, P.; Veeh, R.; Fux, C.; Hu, F.Z.; Quigley, M.; Post, J.C. Role of biofilms in neurosurgical device-related infections. Neurosurg. Rev. 2005, 28, 249–255. [Google Scholar] [CrossRef]
- 89Wiseman, G.M. The hemolysins of Staphylococcus aureus. Bacteriol. Rev. 1975, 39, 317–344. [Google Scholar] [CrossRef] [PubMed]
- 90Darfeuille-Michaud, A.; Aubel, D.; Chauviere, G.; Rich, C.; Bourges, M.; Servin, A.; Joly, B. Adhesion of enterotoxigenic Escherichia coli to the human colon carcinoma cell line caco-2 in culture. Infect. Immun. 1990, 58, 893–902. [Google Scholar] [CrossRef]
- 91Inaba, H.; Nomura, R.; Kato, Y.; Takeuchi, H.; Amano, A.; Asai, F.; Nakano, K.; Lamont, R.J.; Matsumoto-Nakano, M. Adhesion and invasion of gingival epithelial cells by Porphyromonas gulae. PLoS ONE 2019, 14, e0213309. [Google Scholar] [CrossRef] [PubMed]
- 92Inaba, H.; Nomura, R.; Kato, Y.; Takeuchi, H.; Amano, A.; Asai, F.; Nakano, K.; Lamont, R.J.; Matsumoto-Nakano, M. Adhesion and invasion of gingival epithelial cells by Porphyromonas gulae. PLoS ONE 2019, 14, e0213309. [Google Scholar] [CrossRef] [PubMed]
- 93Figueiredo, P.M.; Furumura, M.T.; Santos, A.M.; Sousa, A.C.; Kota, D.J.; Levy, C.E.; Yano, T. Cytotoxic activity of clinical Stenotrophomonas maltophilia. Lett. Appl. Microbiol. 2006, 43, 443–449. [Google Scholar] [CrossRef] [PubMed]