Warum manche Menschen mehr zunehmen als andere, selbst wenn sie sich gleich ernähren
Quelle: Universität Kopenhagen – Naturwissenschaftliche Fakultät.
DDie Gemeinschaft der Darmbakterien, die sogenannte Darmmikrobiota, ist mit nicht weniger als 100 Milliarden pro Gramm Stuhl vergleichbar mit einer ganzen Galaxie in unserem Darm.In einer Gesellschaft, in der das Essen im Überfluss vorhanden ist, kann es für die Gesundheit problematisch sein, wenn einige von uns schon beim Anblick eines Tellers mit Weihnachtsplätzchen zunehmen, während andere mit Genuss essen können und scheinbar kein Gramm zunehmen. Eine Erklärung für dieses Phänomen könnte in der unterschiedlichen Zusammensetzung unserer Darmmikroben liegen. Dies geht aus einer Studie des Instituts für Ernährung, Bewegung und Sport der Universität Kopenhagen hervor.
Um abzuschätzen, wie effizient die Darmmikroben Energie aus der Nahrung gewinnen, untersuchten die Forscherinnen und Forscher die Restenergie im Stuhl von 85 Dänen. Gleichzeitig kartierten sie die Zusammensetzung der Mikroben bei jedem Teilnehmer.
Die Ergebnisse zeigten, dass etwa 40 Prozent der Teilnehmenden zu einer Gruppe gehörten, die im Durchschnitt mehr Energie aus der Nahrung aufnahm als die anderen 60 Prozent. Dabei zeigte sich, dass diejenigen, die am meisten Energie aus der Nahrung gewinnen, im Durchschnitt 10 Prozent mehr wiegen, was einer Zunahme von 9 Kilogramm gleichkam.
„Vielleicht haben wir hier einen Schlüssel gefunden, um zu verstehen, warum manche Menschen mehr zunehmen als andere, obwohl sie nicht mehr oder anders essen. Aber das muss noch weiter erforscht werden“, erklärt Associate Professor Henrik Roager von der Abteilung für Ernährung, Bewegung und Sport an der Universität Kopenhagen.
Zunahme des Risikos von Übergewicht und Adipositas (Fettleibigkeit)
Diese Ergebnisse deuten darauf hin, dass Übergewicht nicht nur eine Frage der Ernährung oder der körperlichen Aktivität ist. Es könnte auch etwas mit der Zusammensetzung der Mikroben zu tun haben, die im Darm eines Menschen leben.
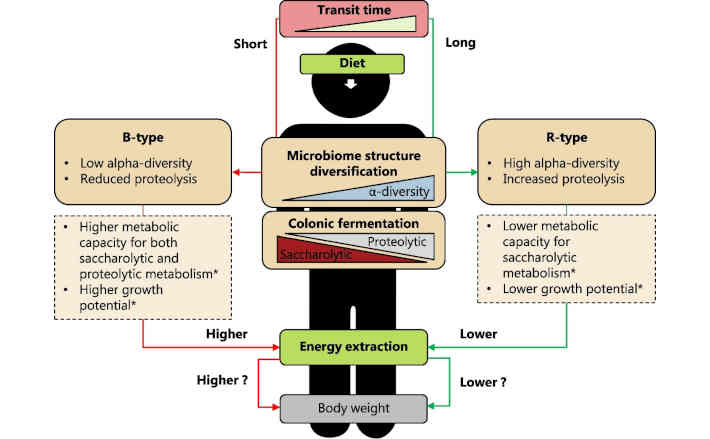
Die Probanden wurden je nach Zusammensetzung ihrer Darmmikroben in drei Gruppen eingeteilt. Die sogenannte B-Zusammensetzung, die von Bacteroides-Bakterien dominiert wird und für die Nährstoffaufnahme effizienter ist, wurde bei 40 Prozent der Teilnehmenden beobachtet.
Die Forschenden vermuten, dass ein Teil der Bevölkerung möglicherweise dadurch „benachteiligt“ ist, dass ihre Darmbakterien bei der Energiegewinnung etwas zu effizient sind. Diese Effizienz könnte dazu führen, dass der menschliche Wirt mehr Kalorien aus der gleichen Menge Nahrung erhält.
Was sich im Laufe der Evolution in Zeiten knapper Nahrungsressourcen als Überlebensvorteil herausgebildet haben mag, hat unter modernen Lebensbedingungen nun oft fatale Nebenwirkungen.
„Die Tatsache, dass unsere Darmbakterien hervorragend darin sind, Energie aus der Nahrung zu gewinnen, ist eigentlich eine gute Sache, denn die bakterielle Verstoffwechselung der Nahrung liefert zusätzliche Energie, zum Beispiel in Form von kurzkettigen Fettsäuren, Molekülen, die unser Körper als zusätzlichen „Brennstoff“ nutzen kann. Wenn wir aber mehr essen als wir verbrennen, kann die zusätzliche Energie, die die Darmbakterien liefern, mit der Zeit das Risiko für Fettleibigkeit erhöhen“, sagt Henrik Roager.
Überraschendes Ergebnis der kurzen Darmpassage
Vom Mund über die Speiseröhre, den Magen, den Zwölffingerdarm, den Dünndarm, den Dickdarm und schließlich den Mastdarm legt die Nahrung, die wir zu uns nehmen, eine Reise von 12 bis 36 Stunden zurück.
Die Forscherinnen und Forscher untersuchten auch, wie lange diese Darmpassage bei allen Teilnehmenden mit ähnlichem Ernährungsverhalten dauerte. Sie stellten die Hypothese auf, dass Menschen mit einer langen Verdauungszeit auch die meisten Nährstoffe aus der Nahrung ziehen. Die Studie zeigte jedoch genau das Gegenteil.
„Wir dachten, dass bei einer längeren Verdauungszeit mehr Energie gewonnen wird. Aber hier sehen wir, dass die Teilnehmenden mit den B-Typ-Darmbakterien, die die meiste Energie extrahieren, auch die schnellste Passage durch das Magen-Darm-System haben, und das hat uns zum Nachdenken gebracht“, sagt Henrik Roager.
Über Darmbakterien
- Jeder Mensch hat eine einzigartige Zusammensetzung von Darmbakterien, die durch Genetik, Umwelt, Lebensstil und Ernährung beeinflusst wird.
- Die Ansammlung von Darmbakterien, die so genannte Darmmikrobiota, ist wie eine ganze Galaxie in unserem Darm, mit erstaunlichen 100 Milliarden von ihnen pro Gramm Stuhl.
- Die Darmbakterien im Dickdarm haben die Aufgabe, Nahrungsbestandteile aufzuspalten, die unsere körpereigenen Verdauungsenzyme nicht verarbeiten können, z. B. Ballaststoffe.
- Menschen können in drei Gruppen eingeteilt werden, je nach Vorhandensein und Häufigkeit von drei Hauptgruppen von Bakterien, die bei den meisten von uns vorkommen: Typ B (Bacteroides), Typ R (Ruminococcaceae) und Typ P (Prevotella).
Bestätigt frühere Studie an Mäusen
Die Studie am Menschen bestätigt frühere Untersuchungen an Mäusen. Diese hatten gezeigt, dass keimfreie Mäuse, die Darmbakterien von fettleibigen Spendern erhielten, bei gleicher Ernährung mehr an Gewicht zunahmen als Mäuse, die Darmbakterien von schlanken Spendern erhielten.
Schon damals vermuteten die Forschenden, dass die unterschiedliche Gewichtszunahme darauf zurückzuführen sein könnte, dass die Darmbakterien fettleibiger Menschen effizienter Energie aus der Nahrung gewinnen. Diese Theorie wird nun durch die neue Studie der Abteilung Ernährung, Bewegung und Sport bestätigt.
„Es ist sehr interessant, dass die Gruppe der Menschen, die weniger Energie im Stuhl hat, im Durchschnitt auch mehr wiegt. Allerdings liefert diese Studie keinen Beweis dafür, dass die beiden Faktoren direkt miteinander zusammenhängen. Wir hoffen, dies in Zukunft genauer untersuchen zu können“, sagt Henrik Roager.
Über die Studie
- Lesen Sie den wissenschaftlichen Artikel: Boekhorst, J., Venlet, N., Procházková, N. et al. Stool energy density is positively correlated to intestinal transit time and related to microbial enterotypes. Microbiome 10, 223 (2022). https://doi.org/10.1186/s40168-022-01418-5
- Sehen Sie sich das Video über die Studie an: Finding connections between the gut microbial community and energy extraction from food
- Der Energiegehalt von Stuhlproben von 85 übergewichtigen dänischen Frauen und Männern wurde untersucht.
- Zu den Teilnehmern gehörten Männer und Frauen im Alter von 22 bis 66 Jahren.
- 40 Prozent der Teilnehmenden fielen in eine besondere Gruppe, die sich durch eine geringere Vielfalt an Darmbakterien und eine schnellere Reisezeit der Nahrung durch ihren Verdauungstrakt auszeichnete.
- Bei dieser Gruppe wurde auch festgestellt, dass sie im Vergleich zu den anderen beiden Gruppen weniger Restenergie im Stuhl hatte, was nicht durch Unterschiede in der gewohnten Ernährung erklärt werden konnte.
- Die Forschenden stellten außerdem fest, dass die Gruppe mit weniger Energie im Stuhl auch mehr wog als die anderen Gruppen.
Kontakt
Zur Ansicht wird JavaScript benötigt!Info-Krümel
Materialien zur Verfügung gestellt durch: Universität Kopenhagen – Fakultät für Naturwissenschaften.
Der Inhalt wurde aus dem Englischen ins Deutsche übersetzt und von der Redaktion überarbeitet.
Tooltips:
Die Texte in eventuell vorhandenen Tooltip-Fenstern wurden von der Redaktion des W3punkt.de bereitgestellt, sie entstammen in der Hauptsache den englisch- und deutschsprachigen Wikipedias.
Wurde der Wissenschaftspublikation entnommen. Die Vertonung des Beitrags erfolgte durch die W3Punkt-Redaktion.
Bildunterschrift:
Vorschlag für eine von der menschlichen Darmmikrobiota abhängige Energiegewinnung. Wir schlagen vor, dass die Transitzeit des Darms die Gemeinschaftsstrukturen des Darmmikrobioms in bevorzugte Gemeinschaftsstrukturen diversifiziert, die von den Enterotypen (B-Typ, Bacteroides; R-Typ, Ruminococcaceae; und P-Typ, Prevotella) erfasst werden, wobei sich der B-Typ und der R-Typ am stärksten in Bezug auf Transitzeit und Alpha-Diversität unterscheiden. Die Enterotypen vom B- und R-Typ unterscheiden sich in der Alpha-Diversität und der Fermentation im Dickdarm, die im Wesentlichen ein Kompromiss zwischen dem saccharolytischen und dem proteolytischen Stoffwechsel ist, was die Gesamteffizienz der Enterotypen bei der Energiegewinnung aus der Nahrung beeinflussen kann. Dies könnte sich möglicherweise in einem unterschiedlichen Körpergewicht niederschlagen. Die Sterne (*) beziehen sich auf die Ergebnisse einer bereits veröffentlichten Arbeit, die zeigt, dass sich die Enterotypen in ihrer Stoffwechselkapazität und ihrem Wachstumspotenzial unterscheiden